河南大学孙旭武&郑州果树所王力荣在《Advanced Science》发文:整合时空多组学揭示桃果中代谢生态位及毛体介导应激适应的遗传基础
- 2026-07-04 10:59:08
 点击蓝字“代谢组metabolome”,轻松关注不迷路
点击蓝字“代谢组metabolome”,轻松关注不迷路

生科云网址:https://www.bioincloud.tech/
编译:微科盟矫情的农民,编辑:微科盟X、江舜尧。
微科盟原创微文,欢迎转发转载。
论文ID
DOI号: 10.1002/advs.202520438
实验设计
结果
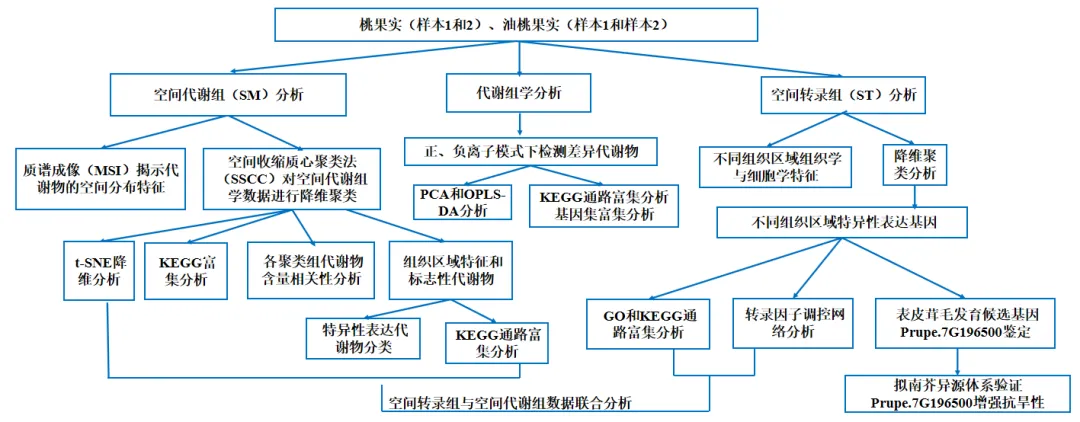
1.桃和油桃果实发育早期的空间代谢组谱图分析
桃和油桃果实在早期发育过程中呈现复杂的空间分布特征。为探究这些代谢物在不同组织区域的表达模式与异质性,研究者利用质谱成像技术(MSI)对授粉7天的未成熟果实进行了空间代谢组谱图特征分析。如图1A所示,油桃与桃果实的整体结构高度相似,桃果实表面存在表皮毛,这是二者最主要的形态学差异。完整的果实结构由果梗、花托、萼片以及果实本体组成。研究者对去除果梗的果实本体进行解剖,可观察到关键内部解剖区域:表皮(外果皮)、中果皮、内果皮、韧皮部、木质部、髓、胚乳、胚、种皮以及果蒂端(图1A)。研究者采用空间收缩质心聚类法(SSCC)对空间代谢组学数据进行降维,获得了果实组织的空间分区信息。图1B、1C表明,在正离子模式(pos)与负离子模式(neg)下检测到的代谢物分布均呈现出规律性空间特征,且与所观察的组织区域高度吻合。借助这些组织切片的注释信息,研究者对经SSCC分析所得的聚类结果完成了类别注释。
接下来,研究者采用t分布随机邻域嵌入算法(t-SNE)对各聚类内的代谢物进行非线性降维分析。t-SNE分布图(图1D、E)清晰显示,不同聚类的代谢物在t-SNE空间内呈空间聚集分布特征,且各聚类占据独立的空间位置。这表明不同组织区域间的代谢物组成存在显著差异。
研究者对不同聚类组代谢物开展京都基因与基因组百科全书(KEGG)通路分析,揭示了各聚类组相关代谢通路的关联性与特异性(图1F、G)。基于各聚类组内代谢物的表达特征与特异性,研究者筛选出每个聚类组表达特异性最高的前10种代谢物,将聚类结果以热图形式可视化,直观呈现不同组织区域代谢物的相对表达水平。这些代谢物的特异表达模式,为解析果实发育过程中代谢物的空间分布规律及其潜在生物学功能提供了重要依据。为进一步探究聚类组间的关联关系,研究者对各聚类组的代谢物含量进行相关性分析。该分析采用皮尔逊相关系数量化变量间线性相关关系,其中红色代表正相关、蓝色代表负相关。相关性分析结果阐释了果实发育进程中不同组织区域间的动态互作规律。
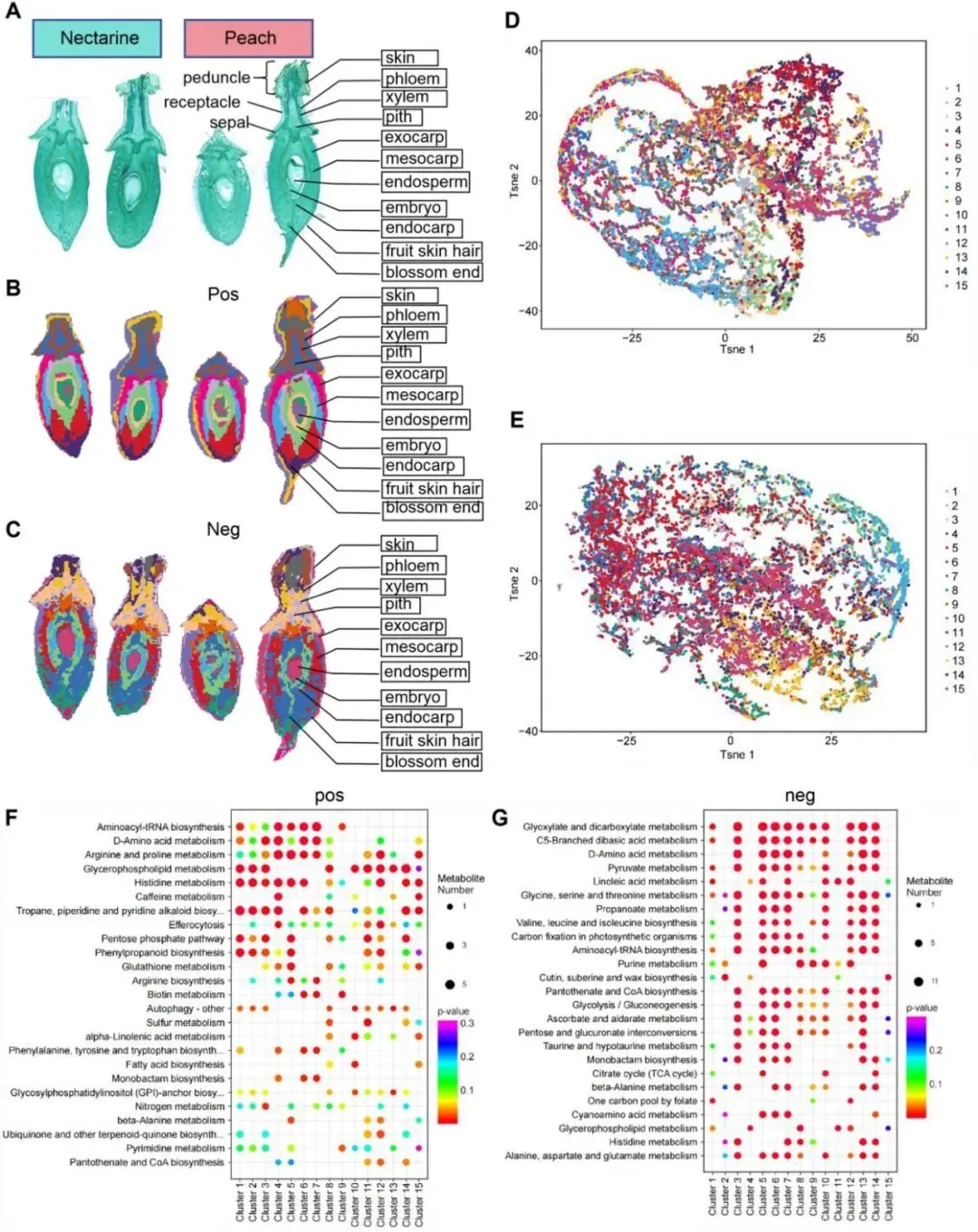
图1 质谱成像(MSI)揭示了桃和油桃中代谢物的空间分布特征。(A)桃和油桃授粉7天幼果的纵向组织切片,解剖标注标示出不同组织区域。(B、C)分别对正离子模式和负离子模式下获取的代谢组数据进行空间收缩质心聚类(SSCC)分析;经降维处理后,不同组织区域的代谢物聚类为15个类群。结果以聚类图谱形式呈现,同一颜色区域代表整体表达谱相似。(D、E)分别对正离子代谢物、负离子代谢物的SSCC聚类结果进行t-SNE降维分析。(F、G)分别对正离子、负离子代谢物经SSCC降维后各类群高表达代谢物机进行KEGG通路富集分析。在图D–G中,为更清晰地展示组织结构与SSCC聚类特征,各类群均以聚类编号标注,而非对应组织区域名称。
2.桃和油桃果实代谢物的空间分布及细胞特征解析
在SSCC降维分析的基础上,研究者筛选出各组织区域特有的代表性代谢物。图2A展示了这些代谢物聚类组在桃果实组织切片中的空间分布特征,而图2B进一步对上述组织区域进行了详细的组织学与细胞学特征分析。果实不同组织区域的细胞类型与形态存在显著差异。具体而言,果柄韧皮部中可见新生韧皮部细胞,木质部中可清晰地观察到新生木质部导管分子,其细胞壁经甲苯胺蓝染色呈蓝色,表明已发生木质化;这类导管分子的形成是果实发育早期为快速生长的果实转运营养物质的关键结构基础。中果皮内层细胞体积更小、排列紧密,细胞密度更高;与之相比,胚乳组织细胞体积明显更大、排列更为疏松,且细胞壁更薄。中果皮内层中还观察到稀疏分布的类维管束结构,由壁厚的长管状细胞构成。研究者对桃果实表皮茸毛的显微观察证实,其起源于表皮细胞,为中空管状的单细胞结构;横切面观察可见典型的中空环状结构。
随后,研究者为每个组织区域筛选了代表性代谢物,并对其空间分布特征进行可视化分析。图2C结果显示,桃果实内代谢物呈现显著的组织特异性空间定位,特定化合物表现出极强的区域分布特异性。研究者对油桃果实开展同步组织学分析,解析了其各组织区域的细胞特征,发现油桃代谢物分布模式与桃高度相似,不同代谢物在特定组织区域中均呈现明显的空间特异性。
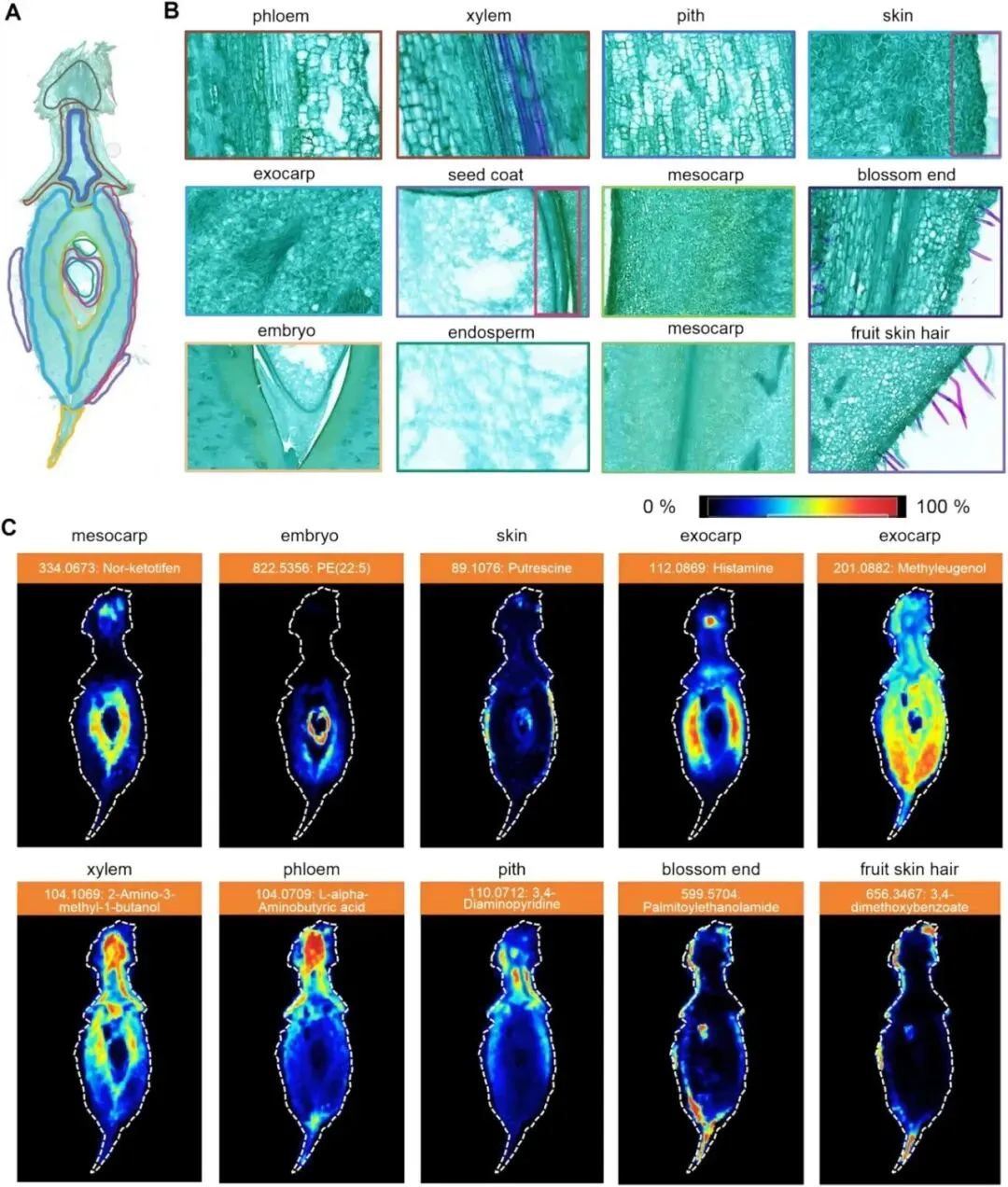
图2桃果实组织区域特征及标志性代谢物表达模式分析。(A,B)桃幼果组织切片不同区域的组织与细胞形态特征。(C)不同组织区域标志性代谢物的表达模式。
3.桃果实中果皮特异性代谢物的代谢谱及通路分析
在代谢物组织特异性分布特征的基础上,本研究对各组织区域的特异性表达代谢物进行分类及KEGG通路富集分析。图3A展示了中果皮中鉴定到的代谢物种类及其相对占比。其中甘油磷酸类占比最高,达8.6%;其次为甘油磷酸甘油类(5.38%)、甘油磷酸肌醇类(5.38%)和甘油磷酸乙醇胺类(4.3%)。三萜类(3.23%)、碳水化合物及其复合物(2.15%)、氨基酸、多肽及其类似物(2.15%)也具有相对较高的占比。其余多种微量代谢物合计占比61.29%,在饼图中统一归为“其他”类别。图3B揭示了中果皮特异性代谢物所参与的代谢通路。多数通路p值偏高,富集显著性较弱;但多条通路表现出统计学显著富集特征。其中甘油磷脂代谢通路富集显著,说明其在桃果实中果皮中可能发挥重要作用;亚油酸代谢通路富集显著性更高(p= 2.51 × 10⁻⁶)。此外,氧化磷酸化、糖基磷脂酰肌醇(GPI)锚生物合成等通路也具有较低的p值,提示具有潜在生物学关联。为验证KEGG富集分析结果,本研究进一步分析了甘油磷脂代谢显著通路中关键中间代谢物的空间表达模式。结果显示(图3C),这类代谢物主要定位于中果皮组织区域。同时,研究者也对内果皮及果实表皮茸毛中特异性富集代谢物的表达模式进行了分析。
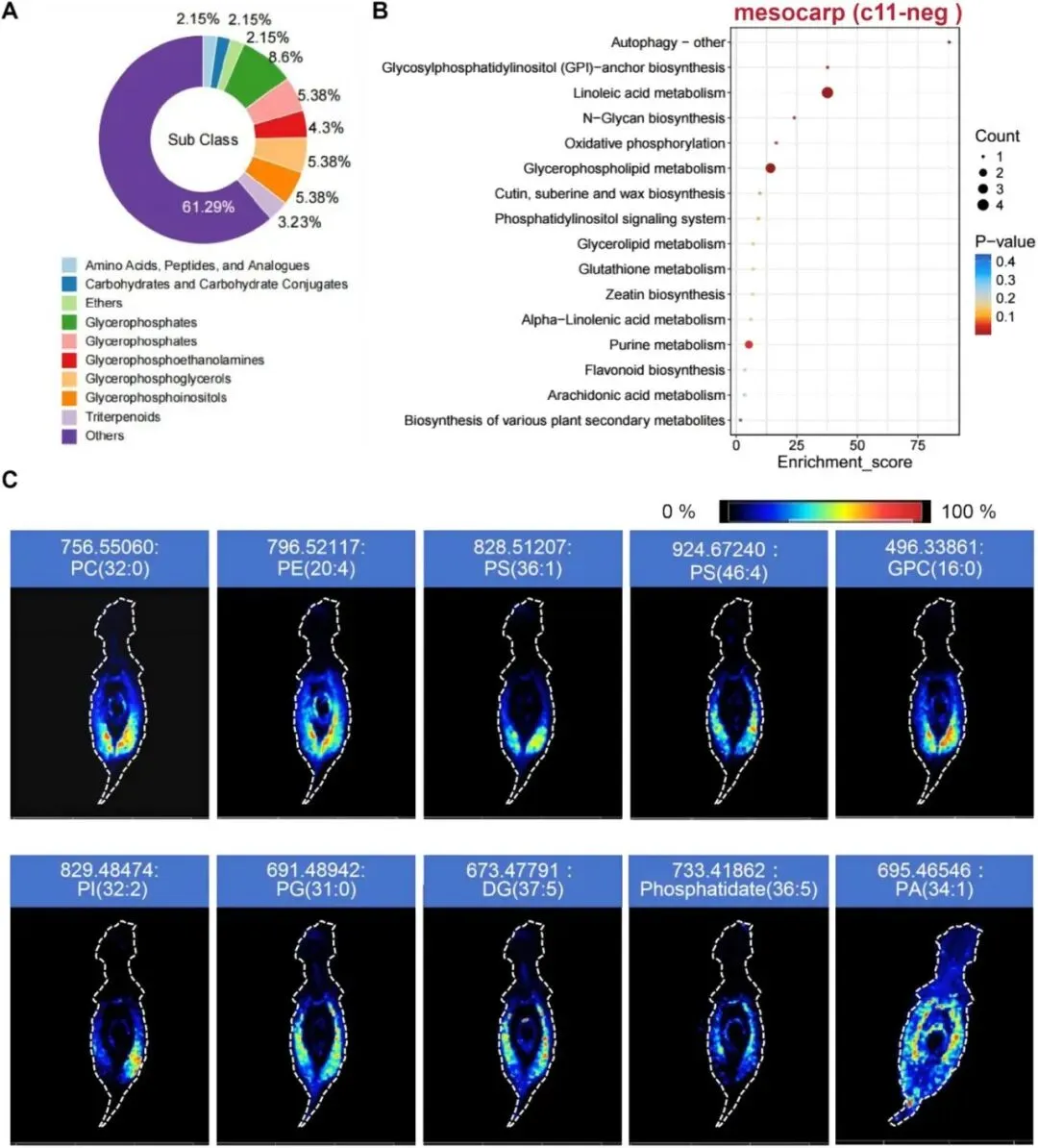
图3 中果皮组织特异性表达代谢物的分类及代谢通路分析。(A)中果皮特异性表达代谢物的类别分布。(B)中果皮特异性代谢物的KEGG通路富集分析。(C)中果皮标志性代谢物的组织特异性表达模式。
4.综合分析揭示油桃与桃的代谢差异
尽管油桃和桃在发育形态上结构差异极小,但二者表皮茸毛发育特征不同,这可能诱导细胞代谢过程产生分化。为验证该假设,研究者对两种果实开展比较代谢组学分析(图4)。研究者对正、负离子模式下检测到的代谢物分别进行无监督主成分分析(PCA)和正交偏最小二乘判别分析(OPLS-DA),结果表明油桃组与桃组间存在显著代谢差异(图4A、B、D、E)。为避免模型过拟合,研究者采用七折交叉验证结合200次响应置换检验(RPT)评估模型稳定性(图4C、F);外部置换检验进一步验证了模型的可靠性。
负离子模式下,相较于桃,油桃中鉴定出285个下调代谢物和334个上调代谢物。采用严格筛选标准(VIP > 1、Benjamini–Hochberg校正后p< 0.05**、**|log2FC| > 1),研究者共筛选得到4个潜在代谢标志物下调、40 个潜在代谢标志物上调(图4G)。正离子模式下,共检测到366个下调代谢物、410个上调代谢物;按照相同筛选标准,获得7个下调标志物与22个上调标志物(图4H)。
KEGG通路富集分析结果显示,两种离子模式下差异代谢物的功能关联存在明显差异:负离子模式差异代谢物主要富集于丙酮酸代谢、乙醛酸与二羧酸代谢、三羧酸循环(TCA循环)以及丙氨酸-天冬氨酸-谷氨酸代谢通路;而正离子模式差异代谢物则与精氨酸和脯氨酸代谢、糖基磷脂酰肌醇(GPI)锚生物合成、甘油磷脂代谢及β- 丙氨酸代谢通路显著相关(图4I、J)。
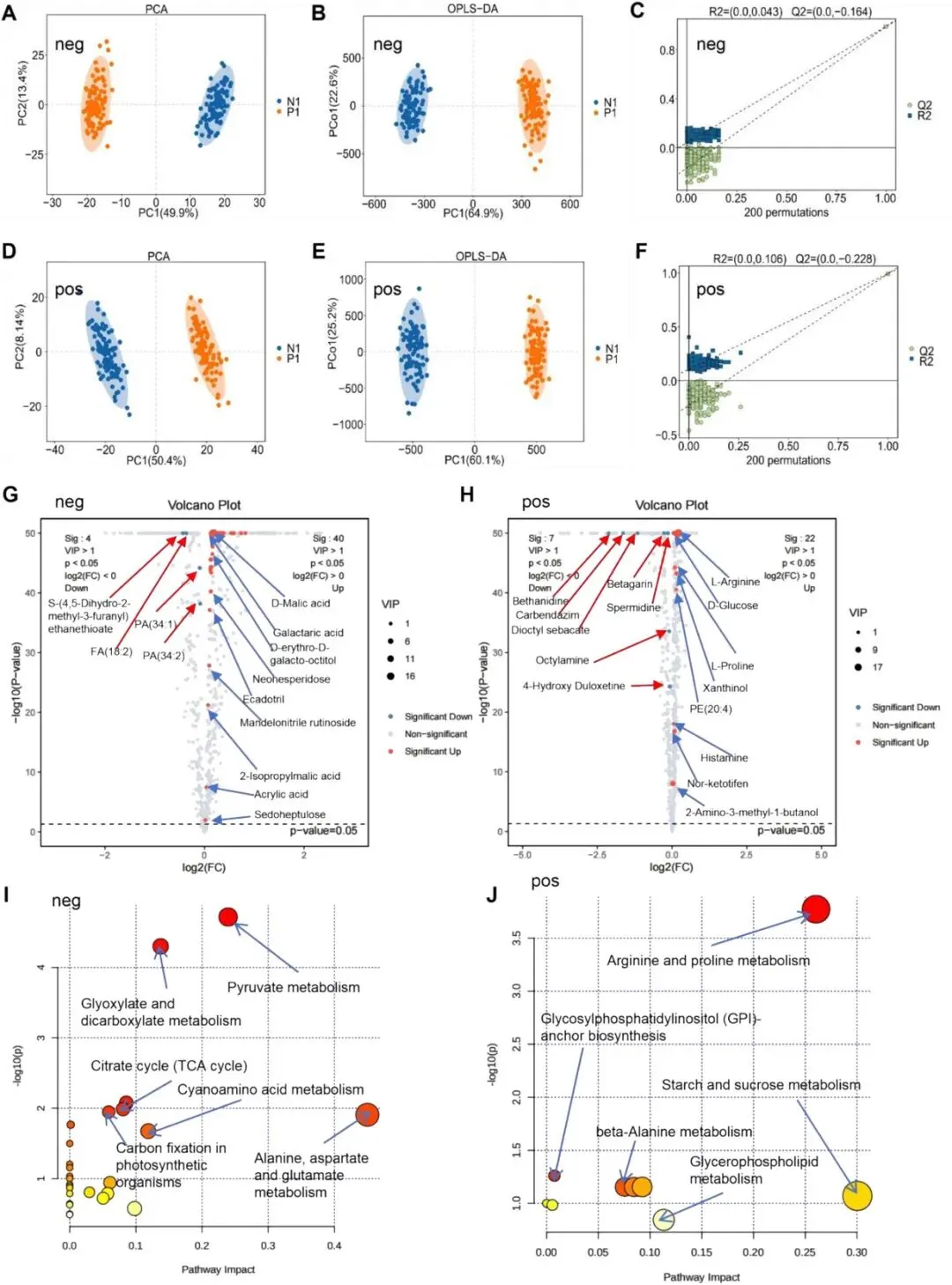
图4 桃与油桃差异代谢物的鉴定与分析。(A)桃和油桃负离子模式下检测代谢物的主成分分析(PCA)。N1(油桃样本1),P1(桃样本1)。(B)桃和油桃负离子模式下检测代谢物的正交偏最小二乘判别分析(OPLS-DA)。N1(油桃样本1),P1(桃样本1)。(C)桃和油桃负离子模式代谢物OPLS-DA模型的响应置换检验。(D)桃和油桃正离子模式下检测代谢物的主成分分析(PCA)。N1(油桃样本1),P1(桃样本1)。(E)桃和油桃正离子模式下检测正离子代谢物的OPLS-DA。N1(油桃样本1),P1(桃样本1)。(F)桃和油桃正离子模式代谢物OPLS-DA模型的响应置换检验。(G)桃和油桃负离子模式下差异积累代谢物的火山图分析。(H)桃和油桃正离子模式下差异积累代谢物的火山图分析。(I)桃和油桃负离子模式差异代谢物的KEGG通路富集分析。(J)桃和油桃正离子模式差异代谢物的KEGG通路富集分析。
5.差异代谢通路的基因集富集分析(GSEA)揭示油桃与桃的代谢差异
为阐明油桃和桃之间的代谢分化特征,本研究对二者代谢物开展基因集富集分析(GSEA)(图5)。研究者对负离子模式数据的分析显示,多条代谢通路存在显著富集差异,通过富集分数(ES)和标准化富集分数(NES)评估,筛选出与品种特异性特征相关的关键代谢通路。甘油磷脂代谢通路表现出明显的差异表达特征(ES = 0.7467,NES = 1.7996;p= 0.0018,FDR = 0.0096;图5A);其核心代谢物磷脂酰胆碱(PC)与磷脂酰乙醇胺(PE)均呈现差异显著(图5A、B),表明二者存在膜脂生物学分化特征,该特征可能对植株生长发育及环境响应具有关键调控作用。戊糖与葡萄糖醛酸互作途径同样存在显著差异(ES = 0.7720,NES = 1.6612;p= 0.0074,FDR = 0.0614;图5A、B)。该通路关键代谢物雌三醇-3-硫酸酯-16-葡萄糖醛酸苷与尿苷二磷酸葡萄糖含量存在显著差异(图5B),表明桃与油桃存在品种特异性的糖代谢与解毒调控机制。此外,嘌呤代谢(ES = 0.6179,NES = 1.5112;p= 0.0399)和半乳糖代谢(ES = 0.7836,NES = 1.4890;p= 0.0200)反映出二者核苷酸与碳水化合物调控通路存在分化(图 5A、B)。在氨基酸代谢通路中,氨酰-tRNA 生物合成(NES = 1.3632;p= 0.1111)与赖氨酸生物合成(NES = 1.3613;p= 0.0875)呈现潜在差异,可能参与调控二者生理性状的分化(图5A)。
研究者对正离子模式代谢物进行GSEA分析,筛选出更多存在品种分化的代谢通路。自噬相关通路呈现显著富集特征(ES = 0.6396,NES = 1.6633;p= 0.0398,FDR = 0.0832),其核心代谢物PE(40:5)、PE(42:10)、PE(36:5)提示油桃与桃的细胞自噬及代谢稳态存在差异。甘油磷脂代谢通路再次表现出强富集效应(ES = 0.4960,NES = 1.6555;p= 0.0133,FDR = 0.0580),涉及PC(37:5)、PE(40:5)、PE(42:10)等代谢物,进一步佐证二者在膜脂代谢层面存在分化。色氨酸代谢(NES = 0.9724;p= 0.4978,FDR = 0.5497)与亚油酸代谢(NES = 1.1342;p= 0.3148,FDR = 0.5521)呈现分化趋势;L-色氨酸、N-甲基血清素、PC (37:5)、PC (35:4)等代谢物可能参与生长调控、抗氧化能力及逆境响应过程。苯丙烷类生物合成通路也表现出特异分化模式(ES = 0.2375,NES = 0.6113;p= 0.9154,FDR = 0.8389),通路中的香草醛与4-羟基肉桂酸等代谢物可能影响果实抗病性与风味品质。
值得注意的是,丙酮酸代谢(负离子模式)与色氨酸代谢(正离子模式,可通过异胡豆苷合成关联萜类吲哚生物碱生物合成)均存在显著差异。两种离子模式下均鉴定到α- 亚麻酸代谢通路的差异代谢物;α- 亚麻酸是茉莉酸(JA)生物合成的前体物质,暗示桃与油桃的茉莉酸信号通路可能存在分化。
为探究代谢的空间调控机制,本研究分析了甘油磷脂代谢通路中关键中间代谢物的组织特异性表达特征。结果表明,这类代谢物存在显著的空间异质性;即使在同一条代谢通路内部,特定核心代谢物的合成与积累也呈现组织空间分区化特征。这揭示了潜在的代谢调控空间格局:代谢物既可在原位合成并发挥局部生理功能,也可通过专一转运机制运输至靶标组织。综上,本研究结果证实:代谢物的合成与空间分配受到精密调控,使其在特定细胞微环境中形成严格的功能特异性,进而维持果实发育进程与生理稳态。
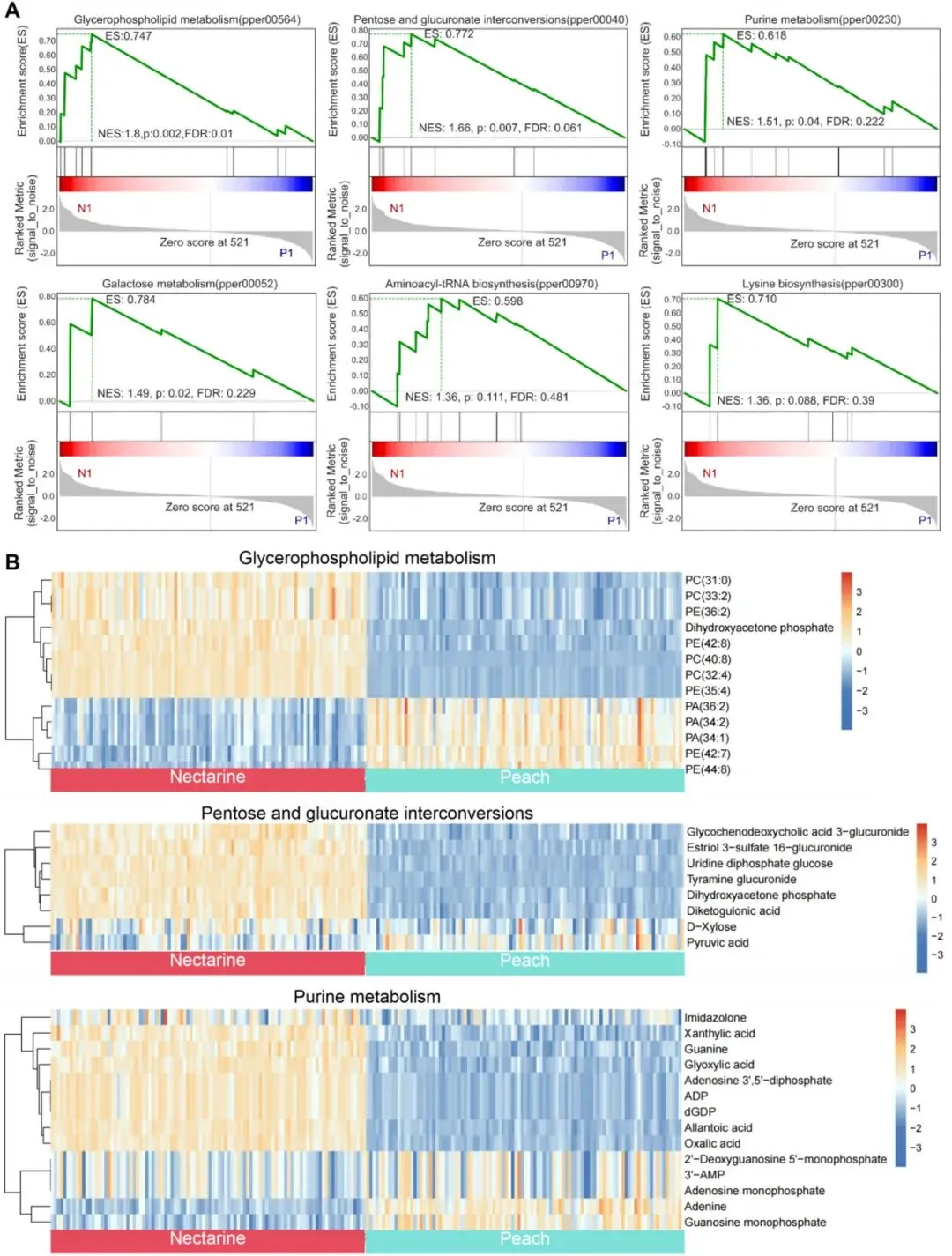
图5 桃与油桃负离子模式差异代谢物的基因集富集分析(GSEA)。(A)桃与油桃负离子模式差异代谢物中代表性代谢通路的GSEA富集曲线图。每张富集图包含四个部分:富集分数(ES)曲线,绿色曲线为动态富集分数,曲线偏离零点的最大峰值对应ES数值;核心富集代谢物:ES>0 时,对富集贡献最大的核心富集代谢物分布于峰值左侧,ES<0时分布于峰值右侧。基因集成员位置:竖线代表基因集内各成员的排序位置。色阶条:由红至蓝的渐变代表排序度量值(如倍数变化、信噪比),红色为正值,蓝色为负值。排序列度量值分布。N1(油桃样本1),P1(桃样本1)。(B)代表性富集代谢通路中相关代谢物表达水平的热图。
6.油桃与桃果实的空间基因表达图谱
在前期空间代谢组学研究明确油桃与桃不同组织间代谢物分布存在显著差异的基础上,本研究开展空间转录组学分析,以解析其内在分子调控机制。试验材料为花后7 天的油桃(油桃样本1、油桃样本2)和桃(桃样本1、桃样本2)样本,各设置2个生物学重复。图6A为桃果实组织切片。经严格质量过滤后,在油桃样本1、油桃样本2、桃样本1、桃样本2中分别鉴定到19 385、19 654、18 817、18 894个基因;各样品分别获得2000、1873、1906、1979个高质量空间位点。各位点平均检测指标如下:基因数分别为3320、4756、2498、2739;UMI数分别为10 756、16 963、8571、9332;线粒体基因占比均处于极低水平。
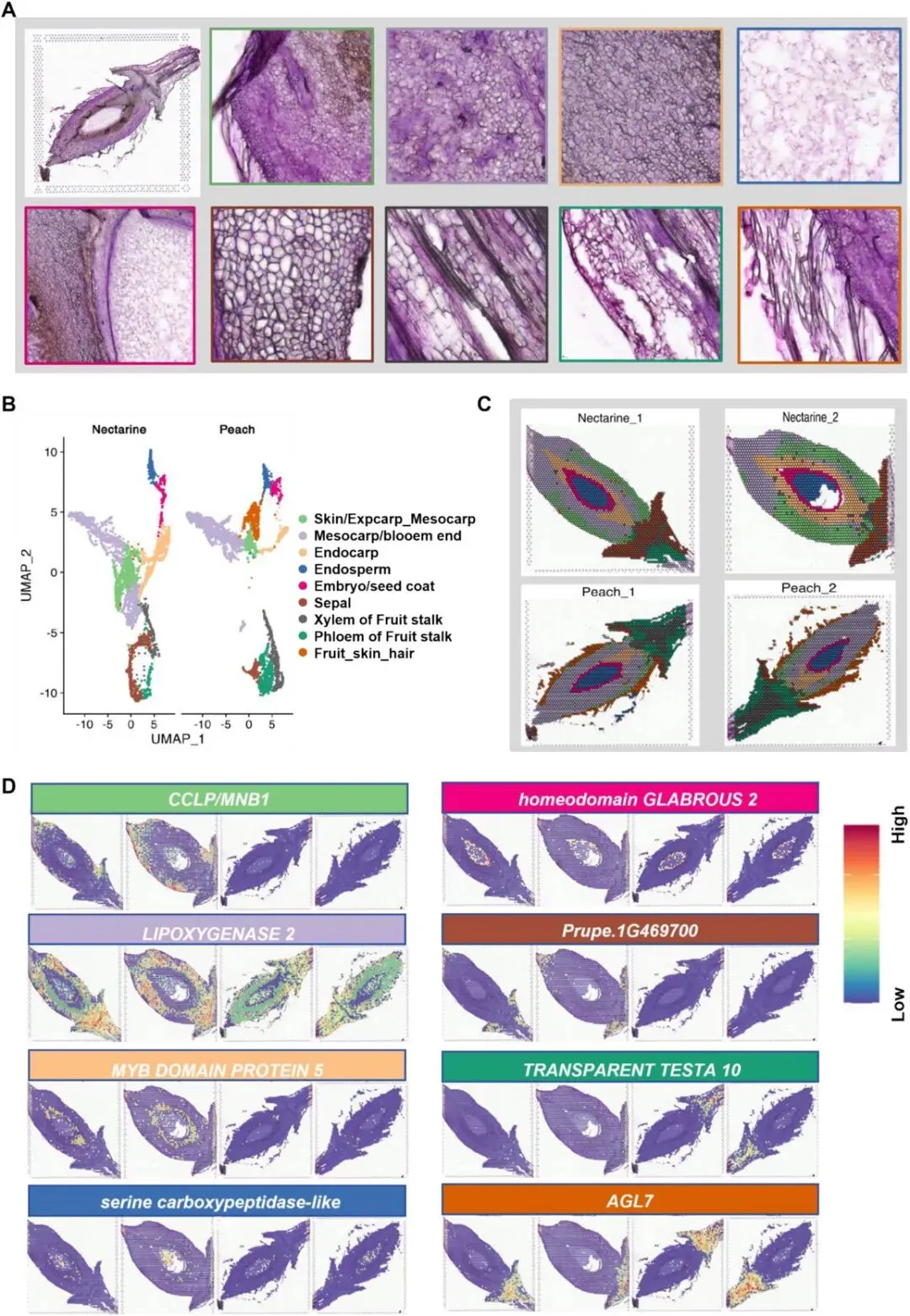
图6 桃和油桃花后7天幼果的空间转录组学分析。(A)桃果实组织切片苏木精-伊红(H&E)染色。方框标注了不同组织区域的组织学与细胞学特征。(B)桃与油桃空间转录组学数据的UMAP降维聚类分析。细胞按9个鉴定聚类组进行着色,并依据对应组织区域完成注释。(C)UMAP聚类结果映射至实体组织切片的空间可视化展示。(D)各聚类组中标志性基因的表达模式。
本研究采用均匀流形逼近与投影算法(UMAP),基于基因表达相似性将所有空间位点聚类为9个不同类群(图6B)。通过空间条形码定位,研究者将各聚类组还原匹配至对应的组织空间位置(图6C)。聚类结果显示,桃与油桃的细胞组成整体相近;仅第9聚类组特异存在于桃样本中(图6B)。值得注意的是,本研究空间转录组数据的空间分辨率有限,无法像代谢组与组织学分析那样将表皮(外果皮)、花萼端、髓部等区域单独区分。因此,表皮与外果皮/中果皮归为一类,花萼端并入中果皮聚类组,髓部区域则归并至木质部聚类组(图6B)。
结合组织切片注释结果,研究者对各聚类组进行如下组织归属定义:聚类组1:表皮/外果皮-中果皮;聚类组2:中果皮/花萼端;聚类组3:内果皮;聚类组4:胚乳;聚类组5:胚/种皮;聚类组6:萼片;聚类组7:果柄木质部;聚类组8:果柄韧皮部;聚类组9:果实表皮茸毛。
虽然各样本间整体测序指标存在一定差异(该现象在空间转录组研究中普遍存在),但核心生物学规律具有高度可重复性。为验证样本重复性,本研究采用两种互补方法开展生物学重复一致性评估。第一种方法:分别计算同一处理下两个生物学重复中、归属相同解剖聚类组的基因表达相关性。各对应组织聚类组(如桃样本1与桃样本2的中果皮)的皮尔逊相关系数均维持较高水平(r> 0.85),表明生物学重复间重复性良好。样本整体指标(单一位点UMI数/基因数)的波动是单细胞测序中常见的技术特征,通常受细胞捕获效率与文库制备效率的微小差异影响;关键在于组织内部相对表达模式保持一致。第二种方法:在全样本层面进一步验证重复性,将每个样本所有位点的表达计数进行合并,构建各生物学重复的假bulk(伪批量)表达谱。研究者对全样本平均表达谱进行皮尔逊相关性分析,结果显示重复间相关性极显著(桃:r= 0.94,p< 2.2e⁻¹⁶;油桃:r= 0.91,p< 2.2e⁻¹⁶),LOESS回归拟合也进一步证实二者呈良好线性相关。该整体水平分析与聚类组层面相关性结果相互印证,证明整体基因表达模式高度可重复。此外,研究者对各组织聚类组的假bulk样本进行层次聚类分析发现:同一组织、同一处理的生物学重复聚为一支,并与其他组织明显分离,进一步说明生物学信号强于技术批次效应。
本研究在各聚类组中鉴定获得功能明确、空间富集的标志基因(图6D、图7):Prupe.4G165500(外果皮/中果皮)为拟南芥甘露糖结合凝集素1(MNBL1,AT1G78830)的同源基因;可在缺铁条件下通过活性氧介导抑制FIT/IRT1/FRO2通路,进而调控铁稳态;同时可通过MYB4-MAN3-甘露糖-MNBL1信号通路,依赖谷胱甘肽介导植物螯合肽合成,增强镉胁迫耐受性,表明该基因参与环境胁迫适应过程。Prupe.2G005300(中果皮)编码脂氧合酶2(LOX2),可催化JA生物合成途径中α- 亚麻酸的氢过氧化反应;通过UVR8-TCP4-LOX2模块介导植株抗UV-B胁迫,在昆虫取食胁迫中参与JA-SA信号交叉调控,并可被NAD+缺失诱导的活性氧激活。该结果与中果皮α- 亚麻酸类代谢物富集特征一致(图6D),表明LOX2在果实防御应答与发育调控中具有重要功能。Prupe.1G405300(内果皮)为MYB5同源基因,可调控种子脂质积累;与TT2协同激活HSFA2以增强耐热性;并可通过与GL2互作调控脂质生物合成通路。Prupe.6G114600(胚乳)编码丝氨酸羧肽酶类蛋白51(SCPL51),为拟南芥AT2G27920的同源基因;功能与水稻OsBISCPL1相似,可在病原菌胁迫与氧化胁迫下激活病程相关基因表达,表明该基因参与胚乳免疫防御过程。Prupe.3G218500(胚/种皮)编码同源异型域蛋白HDG2,为拟南芥AT1G05230同源基因;可直接激活CESA5,参与种子黏液质中纤维素合成,同时与MYB/bHLH/TTG1调控网络协同作用。Prupe.8G047900(木质部)为漆酶LAC15/TT10(AT5G48100)同源基因,可催化木质素聚合与原花青素氧化,对维持细胞壁完整性及维管束功能至关重要。综上,各组织特异基因的空间表达特征表明:果实不同组织中,基因空间定位与其发育及生理功能存在紧密的功能关联。
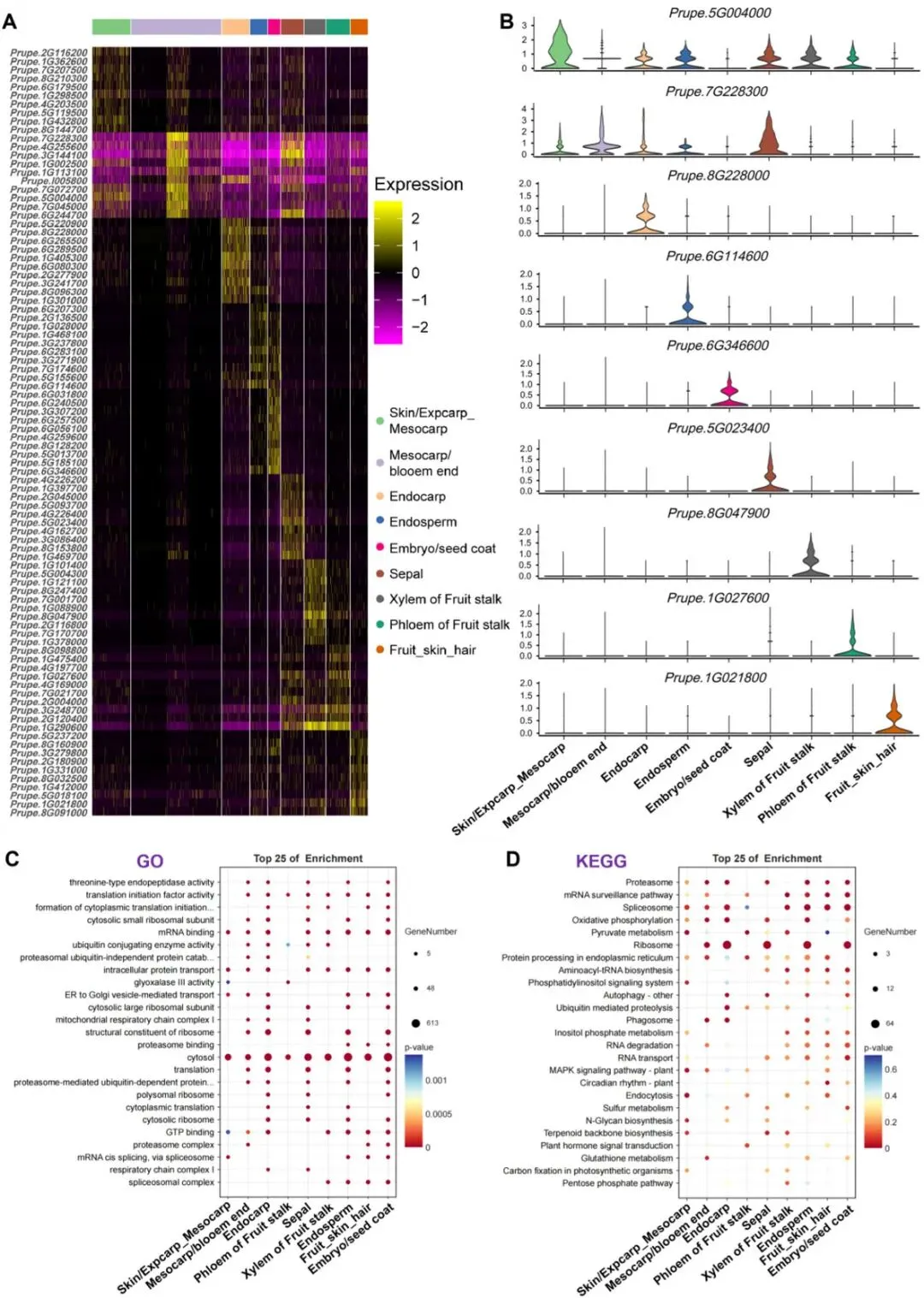
图7 桃与油桃空间限定表达基因的鉴定及功能富集分析。(A)不同组织区域前10个标志基因的表达热图。(B)代表性组织特异基因的表达模式小提琴图。(C)不同组织区域特异性表达基因的基因本体(GO)富集分析。(D)组织特异基因的KEGG通路富集分析。
7.不同组织特异性表达基因的GO与KEGG富集分析
为解析桃果实不同组织的特异性发育与生理功能,本研究鉴定了在各组织中具有空间限定表达模式的基因。图7A以热图展示每个聚类组中表达水平排名前10的组织特异基因;图7B通过小提琴图呈现这些基因在对应组织区域的特异表达特征。研究者进一步基因本体(GO)及KEGG通路富集分析结果显示(图7C、D):脂肪酸代谢、蛋白质降解等基础生物学过程在多个组织中保守存在,说明其在果实生长与生理稳态维持中发挥核心作用。
同时,各组织呈现出独特的功能分化特征:内果皮基因显著富集于吞噬体功能、核糖体活性、胞质生理过程、泛素介导的蛋白水解及呼吸复合体I,有利于维持组织结构完整性与防护功能;外果皮/中果皮基因与代谢调控、系统抗性及液泡组装过程相关,主要参与逆境响应与细胞稳态维持;果实表皮茸毛基因富集于蛋白酶体结合、剪接体复合体及昼夜节律通路,提示其受转录及翻译后调控参与生理进程;中果皮基因还参与碳固定与谷胱甘肽代谢,对果实品质形成及氧化还原平衡至关重要;韧皮部基因调控丙酮酸代谢、乙二醛酶III活性及内质网蛋白质加工过程;木质部基因关联戊糖磷酸途径、萜类骨架生物合成与丙酮酸代谢,体现其营养物质转运分配功能;胚乳/胚基因富集于蛋白质分解代谢、硫代谢及氨酰-tRNA生物合成,是维持蛋白质稳态与胚胎早期发育的关键;萼片基因参与萜类生物合成与蛋白质修复通路,与其花器官保护生理功能相匹配。
研究者补充的KEGG通路分析进一步鉴定出丙酮酸代谢、蛋白酶体功能等在所有组织中均保守存在的通路,凸显其对果实发育的基础性作用(图7D)。值得关注的是,MAPK信号通路成为贯穿多个组织的核心调控枢纽,在整合环境信号与发育程序调控中发挥关键作用。综上,本研究结果表明桃果实各组织间存在精细的功能分化与调控互作关系,为解析其适应性生理机制提供了理论依据。
8.果实表皮茸毛相关标志基因筛选
为丰富桃果实表皮茸毛的标志基因资源,本研究对茸毛富集聚类组9中高表达基因开展空间表达特征分析。结果显示,这些基因在果实发育早期具有显著的时空表达特异性,可作为表皮茸毛发育的分子标志基因。功能注释表明,这些基因在非生物胁迫响应中功能保守:CEL1(Prupe.5G018100)可通过纤维素水解促进细胞壁松弛,参与细胞伸长过程;该基因表达受抑会导致细胞壁褶皱畸形、木质化程度降低、机械强度下降,在表皮茸毛伸长发育中发挥不可或缺的作用。ELIP1作为光保护分子伴侣,可在强光条件下螯合游离叶绿素;基因缺失会加剧光氧化损伤,过表达则可增强植株耐强光能力,推测其对发育中的表皮茸毛具有光保护作用。GATA22可整合细胞分裂素与光信号调控细胞分化,并能依环境背景调控防御反应,可能参与表皮茸毛的起始分化过程。GASA14通过清除活性氧精细调控赤霉素介导的生长进程;功能缺失会抑制叶片扩展、降低胁迫耐受性,过表达则可增强机体解毒能力,表明其在表皮茸毛快速伸长过程中维持氧化还原稳态。MYBD编码MYB结构域转录因子,可通过抑制MYBL2,正向调控光信号与细胞分裂素诱导的花青素积累。
9.油桃与桃果实不同组织差异表达基因的筛选及分析
油桃(果皮光滑发亮)与桃(果皮被覆表皮茸毛)之间的表型分化,反映了果实成熟过程中基因表达差异。为明确表皮茸毛发育差异是否会影响果实整体发育并改变其他组织生理功能,本研究系统鉴定了果实各组织间的差异表达基因(DEGs)(图8A),并进一步开展GO和KEGG富集分析,解析其发育调控分子机制(图8B–E)。
对比分析显示,油桃存在组织特异性的基因下调模式:内果皮中水分转运与营养生长发育相关基因表达下调;外果皮/中果皮中强光响应、脱落酸信号通路及种子萌发相关基因表达下调;维管组织(木质部/韧皮部)中水分转运、碳代谢(如β- 淀粉酶活性)及昆虫胁迫响应相关基因表达下调;种子组织(胚乳/胚)中细胞分裂素生物合成与水通道蛋白活性相关基因表达下调(图8B)。
油桃特异上调基因呈现明显的组织功能差异化富集特征:内果皮富集于类黄酮与原花青素生物合成通路;外果皮/中果皮富集于生物胁迫响应与皂苷生物合成通路;韧皮部富集于光系统II组装与β-半乳糖苷酶活性相关过程;木质部富集于酪氨酸分解代谢、泛素结合及氧化还原过程;胚乳富集于氧化还原代谢;胚富集于胞质核糖体生物发生;萼片则富集于生物胁迫刺激响应通路(图8C)。
KEGG通路分析进一步表明,油桃在多条通路中呈现基因下调:内果皮(丙酮酸代谢)、外果皮/中果皮(植物激素信号通路)、木质部(淀粉与蔗糖代谢、丙酮酸代谢)、胚乳(丙酮酸代谢)以及萼片(激素信号通路、淀粉蔗糖代谢、丙酮酸代谢)(图8D)。上调通路方面,除核糖体通路在各组织普遍富集外,MAPK信号通路与植物激素通路在内果皮、外果皮/中果皮中呈组织特异性上调(图8E)。
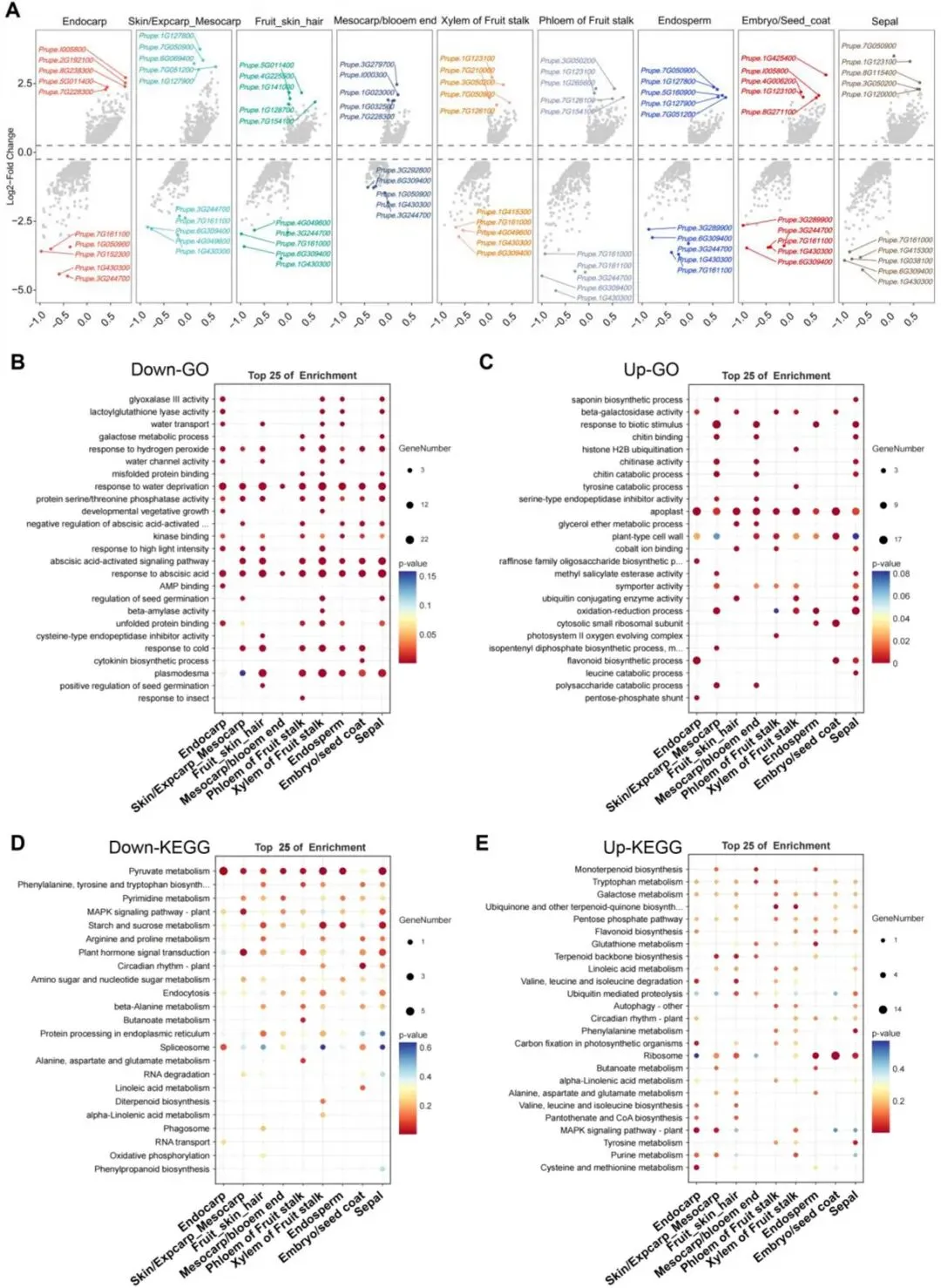
图8 油桃与桃不同组织区域差异表达基因(DEGs)的鉴定及功能富集分析。(A)各组织区域差异表达基因的火山图。每组比较中上调和下调排名前5的基因已突出标注。(B)油桃相较于桃下调差异表达基因的基因本体(GO)富集分析。(C)油桃相较于桃上调差异表达基因的GO富集分析。(D)下调差异表达基因的KEGG通路富集分析。(E)上调差异表达基因的KEGG通路富集分析。
10.油桃与桃差异表达基因相关转录因子调控网络分析
研究者进一步对油桃和桃的差异表达基因(DEGs)开展转录因子调控网络分析,筛选调控差异基因的核心转录因子,并解析转录因子与靶基因间的调控关系。研究者分析共鉴定出9个关键转录因子:3个WRKY家族、3个AP2/ERF-ERF家族、2个bZIP家族以及1个HB-HD-ZIP家族。上述四类转录因子家族均已被证实是介导生物与非生物胁迫响应的重要调控因子。
11.空间组织表达中基因-代谢物关联分析
为探究不同组织和空间维度下基因表达与对应代谢物之间的关联,本研究对空间转录组(ST)与空间代谢组(SM)数据开展联合分析。研究者选取参与桃与油桃代谢物差异积累的多条代表性代谢通路为研究对象,包括:甘油磷脂代谢、丙酮酸代谢、色氨酸代谢、淀粉与蔗糖代谢,并基于上述通路开展基因-代谢物关联分析。通过筛选参与这些代谢过程的功能基因,研究者系统解析其在不同组织中的表达特征。四条代谢通路相关基因在多组织中的表达谱表明,各通路中的代表性基因均具有较强的组织特异表达特征,表明其表达存在明显的时空偏好性。
为进一步阐明上述基因的空间表达特征,本研究分别绘制每条代谢通路中多个代表性基因在组织切片上的空间分布图。同时,对四条通路中的标志性代谢物在桃和油桃样本中的空间积累模式进行可视化分析。通过基因与代谢物空间分布的对比分析发现:各代谢通路内,代谢物与其对应调控基因存在空间共定位特征,为深入探究二者潜在的调控关系提供了空间层面的研究依据。
12.表皮茸毛发育候选基因Prupe.7G196500鉴定
空间代谢组分析结果表明,油桃与桃在异胡豆苷及JA相关代谢物上存在显著差异(图5)。空间代谢组与空间转录组结果均显示,二者在环境胁迫响应与虫害防御方面存在分化特征,推测与油桃表皮无茸毛的表型特征密切相关。为此,本研究对中果皮及表皮茸毛组织表达基因进行差异分析,筛选得到Prupe.7G196500为调控表皮茸毛发育的候选基因(图9A、B)。GO功能富集显示,该基因显著富集于损伤响应与茉莉酸响应条目。外源茉莉酸可促进非洲虎尾草(Chloris gayanaKunth)表皮茸毛的形成;拟南芥相关研究也证实,茉莉酸处理可诱导JAZ蛋白降解,进而激活WD-repeat/bHLH/MYB茸毛形成调控复合体,启动表皮茸毛发育进程。
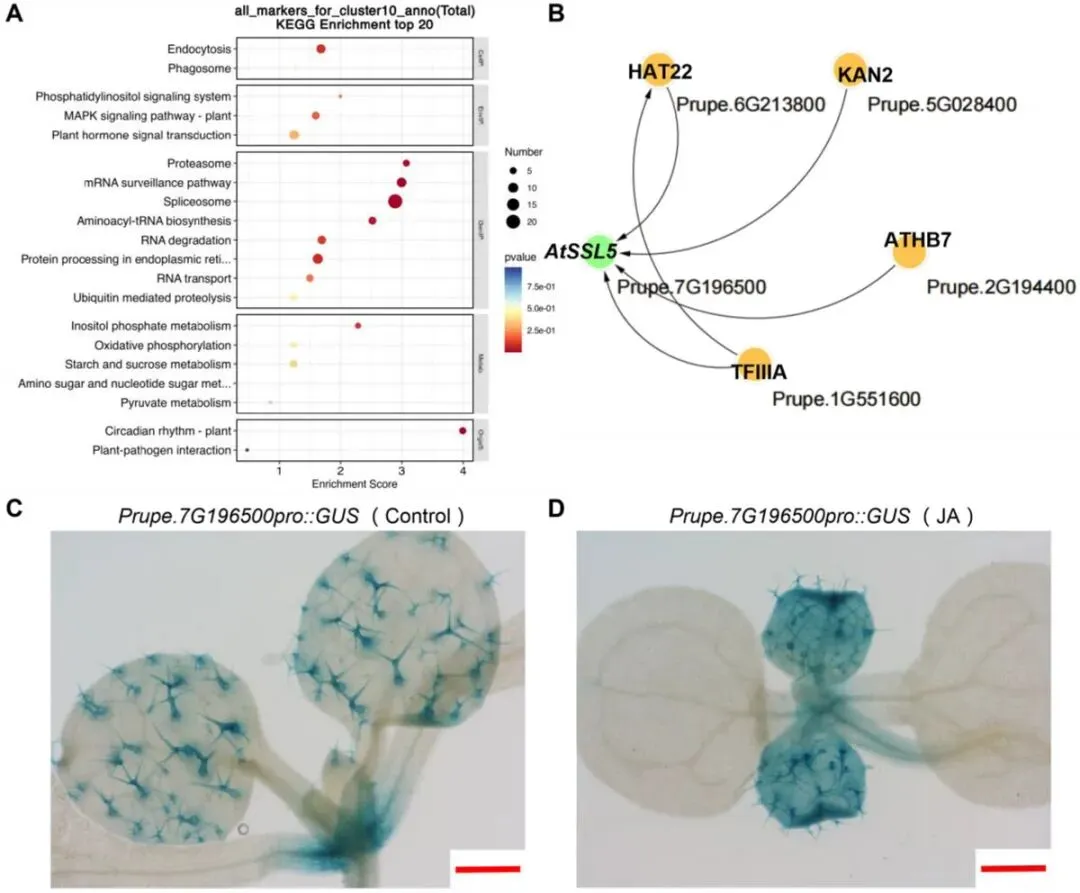
图9 桃果实表皮毛特异基因的鉴定与表达分析。(A)果实表皮毛特异表达基因的KEGG富集分析。(B)表皮毛特异基因的转录调控网络。(C)正常生长条件下,异源表达Prupe.7G196500::GUS的拟南芥植株组织化学GUS染色结果,展示其组织特异性表达模式。(D)茉莉酸处理下转Prupe.7G196500::GUS基因拟南芥植株的GUS染色结果。
Prupe.7G196500与拟南芥异胡豆苷合酶类似蛋白4(AtSSL4)为同源基因,该基因在植物先天免疫中发挥功能。研究者对565份桃种质和179份油桃种质开展表达分析,在Prupe.7G196500的编码区mRNA及启动子区域共鉴定到7处变异位点。关联分析结果表明,位于3'非翻译区的SNP位点(7号染色体:18 561 307 bp)与种质间表皮茸毛密度变异关联最为显著(p= 0.05)。研究者以参考品种‘喀什一号’为材料进行组织特异性表达模式分析,表明该基因主要在果实中高表达。花后7天时,Prupe.7G196500在桃中呈极强的表皮茸毛特异表达,而在油桃中几乎不表达。研究者对‘正白5-2’品种进行果实发育时期表达分析表明,该基因在果实发育早期转录水平达到峰值。研究者对89份种质的茸毛长度统计发现,茸毛长度与基因表达量呈弱相关,且随果实成熟进程茸毛长度逐渐降低。结合基因转录水平与果实生长进程的负向时序关系,研究者认为Prupe.7G196500与表皮茸毛发育呈正相关。
本研究进一步利用拟南芥异源表达体系探究该基因功能。为解析其表达模式,本研究构建了Prupe.7G196500pro::GUS表达载体并转化拟南芥。GUS染色结果显示,该基因在萌发后10天幼苗的表皮茸毛中特异表达(图9C);经40 μmol/LJA处理后染色信号显著增强(图9D),表明该基因受JA诱导表达。正常生长条件下,35S::Prupe.7G196500-1/-2过表达株系的表皮茸毛密度显著高于野生型(WT)。JA处理可进一步提高过表达株系与野生型的茸毛密度,但转基因株系茸毛数量仍显著高于野生型。两种生长条件下,35S::Prupe.7G196500-1/-2株系茸毛数量均显著增加。无JA外源处理时,转基因幼苗已表现出茸毛密度升高的表型,说明Prupe.7G196500过表达可不依赖JA信号通路增强,直接驱动表皮茸毛发育。综上表明:JA可诱导Prupe.7G196500表达并促进茸毛形成,但其调控机制可能与JA信号通路存在差异。
13.拟南芥异源体系验证Prupe.7G196500增强抗旱性
研究者对油桃与桃差异表达基因的GO富集分析显示:油桃整体胁迫敏感性更高,但抗旱能力相较于桃更强(图8B)。已有研究证实,果实表皮茸毛可通过降低表皮水分蒸发调控植株抗旱性;同时鉴于Prupe.7G196500具有促进表皮茸毛发育的功能,本研究提出假说:该基因可通过诱导表皮茸毛形成进而增强植株抗旱性。为验证这一假说,进一步探究了Prupe.7G196500在干旱胁迫应答中的作用。经100 μmol/L甘露醇处理后,GUS染色信号显著增强,表明干旱可诱导Prupe.7G196500表达上调(图10A、B)。正常培养条件下,野生型(WT)与过表达株系幼苗在MS培养基上生长表型无明显差异(图10C);而干旱胁迫处理后,相较于野生型,过表达株系叶片更大、叶色更绿,主根长度也显著更长(图10D、E)。
移栽至营养土的幼苗在正常生长条件下长势无明显差异(图10F)。连续干旱处理14天后,野生型与过表达株系的生长表型仍无统计学差异(图10F)。但持续干旱胁迫21天后,野生型幼苗枯死,而过表达株系可正常存活(图10G)。
上述拟南芥异源体系实验结果证实,Prupe.7G196500参与调控植株干旱胁迫耐受性,可作为桃后续功能研究与育种利用的重要候选基因。
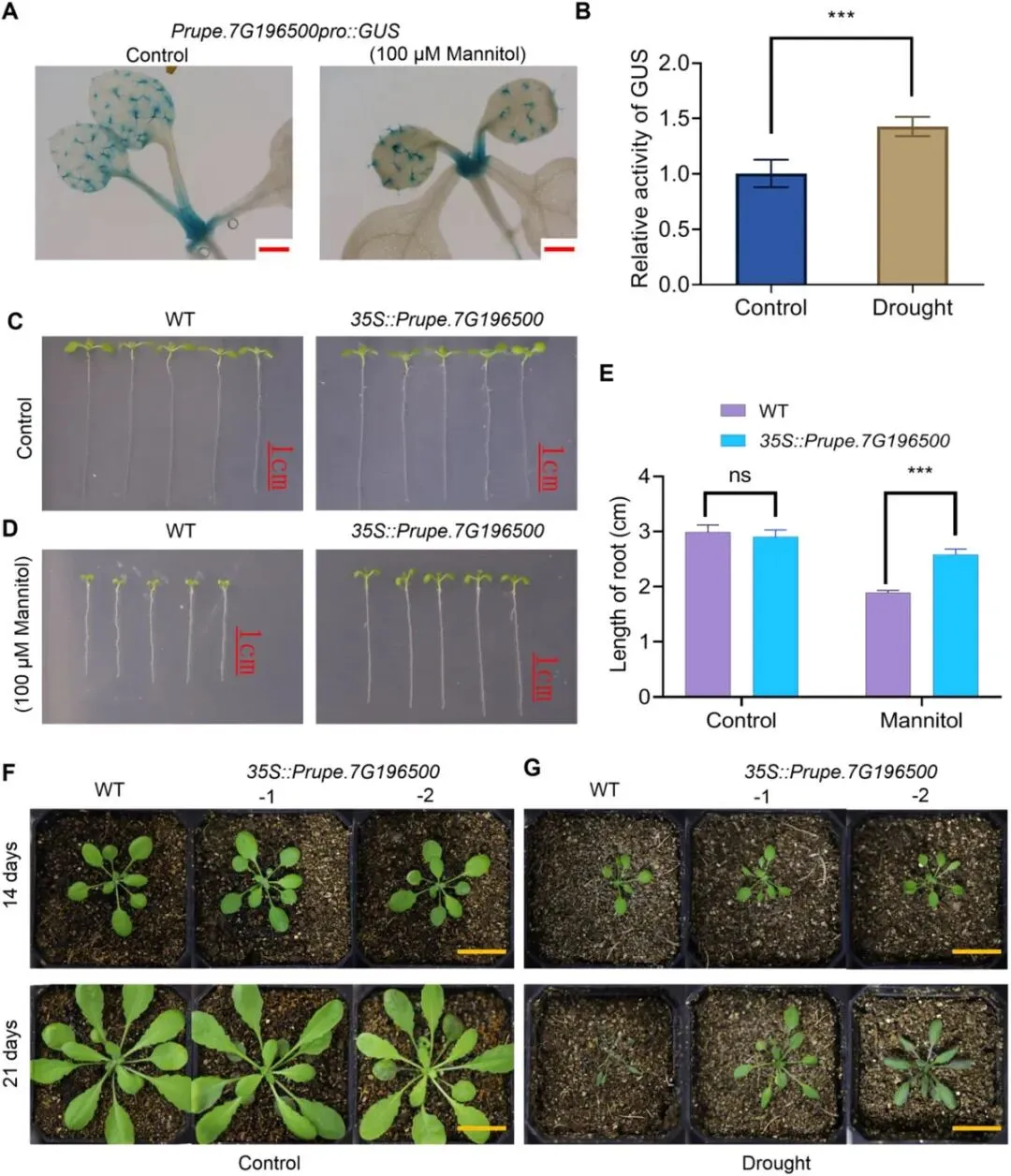
图10 Prupe.7G196500的表达模式与功能分析。(A)对照组与100 μmol/L甘露醇处理下,10日龄转基因幼苗真叶的GUS染色结果。比例尺:500 μm。(B)100μmol/L甘露醇处理后10日龄幼苗真叶的GUS染色信号。比例尺:500 μm。(C)正常条件下,野生型(WT)与35S::Prupe.7G196500过表达株系幼苗在固体MS培养基生长7天的表型。比例尺:1 cm。(D)添加100 μmol/L甘露醇的MS培养基中,野生型与过表达株系幼苗生长7天的表型。比例尺:1 cm。(E)野生型与35S::Prupe.7G196500过表达株系幼苗根长统计分析。***p< 0.001;采用单因素方差分析与野生型比较。(F)正常条件下,野生型与过表达株系幼苗在营养土中生长14天的表型。比例尺:2 cm。(G)干旱胁迫处理下,野生型与过表达株系幼苗在营养土中生长14天的表型。比例尺:2 cm。
讨论
1.果实发育时空图谱:桃的代谢分区与进化创新
本研究整合空间代谢组(SMI)与空间转录组(ST)技术,首次在蔷薇科构建了桃果实早期发育的多维空间图谱。质谱成像(MSI)技术可实现桃与油桃果实组织内代谢物特异性空间分布的可视化,弥补了传统质谱方法缺乏空间定位信息的短板。该技术对于解析果实发育、胁迫应答及分子功能等植物生理过程具有不可替代的重要作用。通过获取代谢物原位时空分布特征,质谱成像能够揭示代谢物在细胞及亚组织结构(如外果皮、中果皮、种区)中的积累模式,有助于阐释代谢物生物合成通路与调控机制,以及代谢物动态变化对果实早期发育的调控作用。油桃是桃表皮茸毛缺失的遗传变异类型。利用质谱成像对比二者果实发育早期代谢物的空间差异(如茸毛相关化合物的空间分布),可为揭示其生理适应机制提供新视角;例如阐明表皮区域代谢物分布如何决定果实果皮光滑度及抗逆性特征。
本研究突破了传统的整块组织分析范式,揭示了代谢物与转录本在不同解剖结构域中的动态分配及如何协同调控三大相互关联的生物学进程:组织功能特化、代谢权衡驱动的物种分化,以及发育调控基因被借用实现胁迫适应性。本文将这些研究见解整合归纳,置于植物空间生物学与果实演化的宏观研究背景下进行阐释。
2.空间代谢分区奠定组织形态建成基础
基于质谱成像构建的代谢空间图谱揭示了果实内部显著的空间异质性,其中甘油磷脂代谢与亚油酸代谢通路主要定位于中果皮(图3B)。关键中间代谢物如磷脂酰胆碱PC(37:5)、磷脂酰乙醇胺PE(42:7)、PE(42:8)富集于外果皮及表皮区域,呈现典型的空间代谢分区特征,即代谢通路通量在物理空间上被限定于特定组织分区内。在幼果期(YF),甘油磷脂代谢相关基因与代谢物均呈现动态变化;其中溶血磷脂(LPLs)等甘油磷脂类代谢物在幼果期相对丰度显著高于成熟期(MF),表达水平明显上调。这表明幼果期是甘油磷脂生物合成的活跃阶段。
在果实发育早期(如番茄花后5天内),脂质代谢相关基因表达上调,并与甘油磷脂组分变化显著相关。具体而言,SlKLUH等基因低表达可促进甘油脂、磷脂等脂质代谢通路基因上调,这表明果实发育起始阶段存在甘油磷脂代谢的转录激活过程。
甘油磷脂代谢相关蛋白(如参与甘油脂代谢的关键酶)在果实早期发育过程中也发生明显丰度变化,且在发育初期表达水平更高。蛋白质组学分析表明,这类蛋白的丰度差异可能与幼果期特有的代谢需求相匹配。
甘油磷脂代谢通路(如甘油脂代谢)是调控果实发育早期脂质积累的关键通路,为果实油脂合成与能量贮存提供物质基础。幼果期糖代谢对脂质生物合成的调控作用尤为显著;甘油磷脂可作为中间代谢物,助力脂肪酸合成与膜脂形成。此外,甘油磷脂代谢还可与磷脂酰肌醇生物合成等其他通路发生交联互作,通过转录调控因子调控脂质积累,进而维系果实早期生长发育。甘油磷脂作为磷脂的主要组成成分,参与膜脂组装过程(如磷脂酰乙醇胺与溶血磷脂的合成),支撑幼果期细胞快速分裂与膨大。该过程既维持细胞结构完整性,也为后续果实成熟等发育阶段奠定生理基础。在腰果、油棕等物种的发育进程中,甘油磷脂代谢的改变还可影响膜流动性,并调控磷脂酰肌醇等信号分子的生成,进而参与生长发育的调控过程。
幼果发育阶段,甘油磷脂代谢可与甘油脂代谢、脂肪酸代谢等通路协同作用,维持机体代谢稳态。转录组与代谢组联合分析表明,甘油磷脂代谢可作为果实发育早期的核心调控枢纽,调控脂质代谢网络的动态平衡,包括脂质降解与生物合成之间的代谢权衡。这种多通路协同调控可优化物质资源分配,进而促进果实生长发育。
转录组空间聚类进一步解析了发育中果实内部的功能模块性。与此结果相一致,LOX2(聚类组2/中果皮)与α-亚麻酸代谢相关代谢物存在空间共定位(图6D),证实该基因具有催化α-亚麻酸氢过氧化的生物学功能。同样,Prupe.4G165500/MNBL1(聚类组1/外果皮)可通过抑制ROS-FIT通路调控铁离子清除过程,这也解释了其为何与抗坏血酸-谷胱甘肽循环代谢物呈现共富集特征(图6D)。桃特有聚类组9中富集ELIP1与GASA14基因,表明光信号与活性氧通路可能参与调控表皮茸毛发育。
3 桃与油桃分化中的补偿性适应机制
油桃的代谢谱发生改变,核心表现为α-亚麻酸代谢通量增强(该物质是茉莉酸的前体,图8E),同时丙酮酸脱氢酶旁路途径的碳分流出现差异分化(图8D)。
这种代谢调控体现了一种进化权衡:油桃因表皮茸毛退化缺失物理屏障,进而导致水分转运与脱落酸信号通路相关基因整体下调(图8B)。与此同时,油桃启动化学抗性通路进行补偿性上调:一方面类黄酮生物合成通路增强(图8C),已有研究表明类黄酮可清除紫外胁迫诱导产生的活性氧;另一方面自噬通路被激活(图8C),可在胁迫条件下降解并回收受损细胞器,提升植株抗逆适应能力。
JA信号通路在表皮茸毛发育与胁迫适应过程中均发挥关键调控作用,成为连接两大生物学过程的分子机制纽带。本研究多方面证据表明,桃与油桃之间存在茉莉酸介导的调控分化:第一,GSEA显示,作为茉莉酸合成前体的α- 亚麻酸代谢通路在两组间存在显著富集差异;第二,表皮茸毛特异标记基因Prupe.7G196500在拟南芥异源实验中表现出明显的茉莉酸诱导表达特性(图9D);第三,本研究鉴定到9个胁迫响应核心转录因子,其中多个为已知的JA通路调控因子:包括PpWRKY75(与拟南芥AtWRKY75同源),该基因可通过调控茉莉酸合成介导植株对坏死营养型真菌的抗性;以及PpERF1(与拟南芥AtERF1同源),为茉莉酸/乙烯信号通路的上游重要组分。
在烟草中,NtHD9与NtHD12双突变体因JA信号通路缺陷,无法形成长腺毛表皮茸毛,且外源施加茉莉酸甲酯(MeJA)也无法恢复其表型。与之类似,拟南芥SVB和SVBL双突变体表现出生长受阻与表皮茸毛发育异常,表明其激素调控网络发生紊乱。
本研究在油桃中观察到的补偿性代谢调控现象,与其他物种已有研究结论高度吻合。例如,黄瓜无毛突变体mict(csgl1/cstbh)的转录组与代谢组联合分析表明:表皮茸毛缺失会导致类黄酮代谢和角质层代谢相关基因显著下调,同时类黄酮、脂质及角质层组分含量发生大幅改变。这直接证实,表皮茸毛缺失会显著影响上述关键代谢通路的活性。与之相一致,拟南芥查尔酮异构酶1缺陷突变体无花青素(af)因缺乏类黄酮,进一步造成腺毛中萜类生物合成相关基因转录水平与酶活性下降。多项研究也进一步佐证了表皮茸毛在植物防御中的重要作用:表皮茸毛发育与角质层形成存在遗传互作,二者协同参与植物防御策略的构建。棉花中GhHD1A通过调控GhGIR1D的表达影响表皮茸毛起始分化;在ghhd1a突变体背景中沉默GhGIR1D可恢复野生型表型,说明GhGIR1D存在负反馈调控或独立调控机制。推测该调控过程可能由次生代谢物等特定代谢物的积累所介导。
表皮茸毛缺失带来的另一重要生物学效应,是植株对生物胁迫的敏感性显著升高。番茄自然突变体tm即为典型例证:其表皮茸毛形态发生改变(茸毛隆起、基部变窄),直接导致植株对蚜虫的抗性显著下降。同样,茶树表皮茸毛的代谢组分析证实,茸毛合成的风味及防御相关代谢物对植物自身防护至关重要。另有研究提出,腺毛的光合作用可为其自身高水平代谢通量提供物质与能量支撑。部分突变体中初级代谢(如糖代谢)发生紊乱,还会间接干扰次生代谢物的合成;拟南芥低钾响应突变体中糖代谢与水杨酸相关代谢物大量积累,即是典型佐证。
调控网络分析进一步筛选得到9个胁迫响应转录因子,参与调控植株逆境应答与环境适应性。其中包含3个WRKY家族转录因子:Prupe.3G098100(PpWRKY40)、Prupe.6G230600(PpWRKY7)和Prupe.1G223200(PpWRKY75)。Prupe.3G098100与拟南芥AtWRKY40同源,该基因可与WRKY18/60协同调控脱落酸信号与非生物胁迫应答。此外,拟南芥WRKY40与WRKY18可共同抑制flg22诱导的防御相关基因表达,避免植物防御反应过度激活。Prupe.6G230600同源于拟南芥AtWRKY7,是丁香假单胞菌防御反应的负调控因子;而Prupe.1G223200对应拟南芥AtWRKY75,通过调控JA合成介导植株对坏死营养型真菌的抗性。
AP2/ERF-ERF家族转录因子包括:Prupe.2G272400(PpERF105)、Prupe.1G037900(PpERF1)和Prupe.1G513600(PpRAP2.4)。Prupe.2G272400与拟南芥AtERF105同源,参与调控低温胁迫适应性及对丁香假单胞菌的抗病反应。Prupe.1G037900同源于拟南芥乙烯响应因子1(ERF1),是茉莉酸/乙烯信号通路的上游关键组分,参与病原菌抗性与盐胁迫应答。Prupe.1G513600与拟南芥AtRAP2.4同源,可响应盐胁迫与干旱胁迫,并能在干旱条件下激活角质蜡质生物合成。
本研究还鉴定到bZIP家族转录因子Prupe.1G434500(PpABF2)与Prupe.2G182800(PpGBF3)。Prupe.1G434500与拟南芥AtABF2同源,在营养生长阶段位于SRK2D/E/I下游,介导脱落酸调控的渗透胁迫应答;同时可在干旱脱水胁迫下与ANAC096发生互作。Prupe.2G182800同源于拟南芥AtGBF3,是调控植株抗旱性的关键正向诱导因子。Prupe.6G193400编码HB-HD-ZIP家族转录因子,与拟南芥AtHB6同源,推测参与脱落酸信号通路调控。以上结果表明,桃与油桃存在明显的发育进程分化;WRKY、AP2/ERF-ERF、bZIP、HB-HD-ZIP四类转录因子在二者逆境应答机制差异中发挥核心调控作用。
4 Prupe.7G196500兼具发育与非生物抗逆双重整合调控功能
本研究鉴定得到Prupe.7G196500(拟南芥SSL4的功能同源基因),是桃中一个全新的双功能调控候选基因,该基因可整合表皮发育与干旱抗逆两大生物学过程。该基因符合双功能调控因子的特征:具有表皮茸毛特异表达模式(图10A),且受JA显著诱导(图9D),位于茸毛起始核心调控模块PpMYB25-PpMYB26的下游。为验证其发育调控功能,本研究在拟南芥中异源过表达Prupe.7G196500,结果发现表皮茸毛密度提升1.2倍。该调控效应并不依赖异源体系中JA信号通路的放大作用,表明其在植物表皮建成调控网络中具有保守功能,尤其可能作用于GL3-EGL3转录复合物通路。但要最终确证其在桃体内作为核心调控因子的功能,仍需后续在桃本生遗传背景下开展功能缺失实验,例如利用CRISPR/Cas9基因敲除或RNAi沉默技术进行验证。
除调控发育功能外,过表达Prupe.7G196500的转基因株系可耐受21天土壤干旱胁迫并正常存活,而野生型植株则枯死死亡(图10G)。结合该基因受茉莉酸诱导且表皮茸毛特异表达的特征,足以证实Prupe.7G196500是整合生长发育与胁迫适应的分子调控枢纽。尽管上述结论来自异源体系验证,但表皮茸毛(叶毛)本就是旱生植物(如柠条)吸收大气水分的关键结构。表皮茸毛密度增加,可帮助植株在干旱条件下维持叶片水势、叶片水力导度(Kleaf)与气体交换能力;同时推迟引发叶片水力导度丧失50%的临界水势出现时间,并在一定程度上维持光系统功能稳定。与之相反,同属近缘但无表皮茸毛的物种(如小叶锦鸡儿)则不具备上述抗旱生理优势。
表皮茸毛可通过吸收露水缓解土壤水分亏缺,减轻干旱胁迫对叶片水力系统的损伤,从而维持植株整体水分平衡。在番茄中,表皮茸毛/气孔比值(T/S)与内在水分利用效率(WUEi)、生物量水分利用效率(WUEb)均呈正相关。这表明表皮茸毛与气孔共同在番茄抗旱性中发挥重要作用,也为培育高水分利用效率主栽作物提供了新的育种思路。
此外,表皮茸毛能够富集镉等重金属离子,通过固持有毒金属离子减轻重金属胁迫伤害,还可间接增强植株对虫害的抵御能力。腺毛可合成萜类、生物碱等特化防御代谢物,直接驱避植食性昆虫;且腺毛密度通常与植株抗虫性呈正相关。本研究填补了一项关键研究空白:阐明多年生木本植物如何在水分亏缺条件下,同时实现体表结构(如依靠茸毛调控微环境)与细胞逆境应答的协同优化。本研究鉴定到的Prupe.7G196500是JA响应、茸毛特异表达基因,且在异源体系中可赋予植株抗旱性,为激素信号通路与植株物理防御、代谢适应之间建立了分子机制关联。从应用价值来看,Prupe.7G196500可作为培育气候韧性李属果树品种的核心候选基因,能够兼顾表皮茸毛介导的结构防御,以及渗透调节物质、活性氧稳态调控介导的代谢保护。
结论
本研究阐明了果实生物学的一项基础调控规律:代谢程序与转录程序的空间分异,驱动不同组织结构域的功能特化。本研究取得多项核心进展:证实代谢分区是植物资源优化分配的一种进化策略,典型特征为甘油磷脂合成通路在中果皮特异性富集。同时,揭示了油桃通过物理屏障(表皮茸毛建成)与生化防御系统(类黄酮、自噬通路等)之间的代谢权衡,形成补偿性适应机制。此外,本研究发现茉莉酸响应型发育基因Prupe.7G196500存在功能适应性演化,可同时作为非生物抗逆候选基因,构建了激素信号— 表皮茸毛发育 — 干旱适应三者关联的分子调控框架。综上,本研究构建的果实时空空间图谱,既可为果树组织工程筛选标记基因等研究提供基础资源,也为解析气候变化背景下多年生作物的发育可塑性奠定了理论范式与研究体系。
原文链接:https://doi.org/10.1002/advs.202520438
----------微科盟精彩文章----------
1 Phytomedicine│成都中医药大学:空间代谢组学结合质谱成像揭示了白芷在偏头痛大鼠中的肠-脑轴机制
2 科研 | 福建农大:利用UPLC-QTOFMS、分子特征网络和DESI-MSI对茶树中皂苷进行空间成像和代谢分析(国人佳作)
如果需要原文pdf,请扫描文末二维码



获取此文献原文PDF、申请加入学术群,联系您所添加的任一微科盟组学老师即可,如未添加过微科盟组学老师,请联系微生态老师10,无需重复添加。
请关注下方名片,了解更多代谢组学知识

