JAR | 郑州烟草研究院等:WRKY6与WRKY23调控烟草对丁香假单胞菌侵染的时空防御机制:多组学整合分析
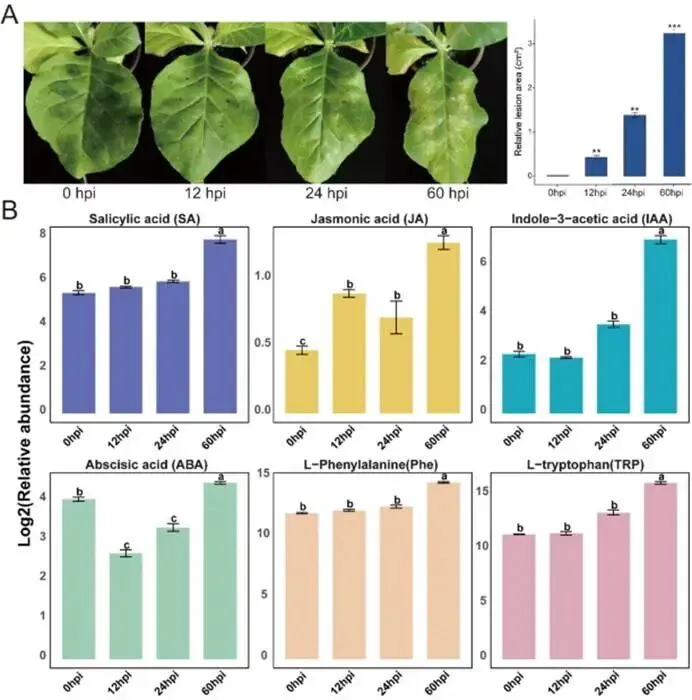
引言:丁香假单胞杆菌烟草致病变种(Pseudomonas syringae pv. tabaci)是一种革兰氏阴性细菌病原体,可引起毁灭性的烟草野火病,在全球范围内造成重大经济损失。尽管其致病性已有充分记载,但烟草在应对该病菌侵染过程中的动态防御机制仍未得到充分阐明。研究目的:本研究旨在通过多组学整合分析,解析烟草抵御丁香假单胞菌(P. syringae)侵染的阶段性防御机制,重点阐明转录重编程与代谢重塑之间的时空协调关系,并对关键调控模块进行功能验证。研究方法:本研究整合了时间序列转录组学和代谢组学数据,以重建动态响应模式。通过采用时序基因共表达网络分析和加权基因共表达网络分析,探究了阶段特异性调控模块。此外,通过创制转基因株系,验证了转录因子WRKY6和WRKY23在抗病性中的作用。研究结果:在侵染早期(12-24小时),烟草优先激活胁迫信号与植物激素通路(水杨酸/茉莉酸),而在侵染后期(48-72小时)则转向细胞稳态调控。WRKY、ERF和NAC转录因子家族协调了阶段特异性的基因表达。值得注意的是,WRKY6和WRKY23发挥负调控作用,其基因沉默可导致病斑面积减少42-58%,病原菌载量降低3.2-4.5倍。代谢组学分析揭示了苯丙烷类代谢途径的持续激活,该途径特异性调控L-苯丙氨酸稳态及其防御性衍生物黄嘌呤核苷的生物合成。此外,研究还鉴定出涉及鞘脂代谢、光响应以及激素信号串扰的核心调控模块。结论:本研究表明,WRKY介导的转录重编程通过协调植物激素信号、鞘脂动态和光响应,在时空上调控次生代谢产物的生成。所鉴定的WRKY6和WRKY23调控模块为设计工程化抗病性提供了分子框架,而本研究提出的两阶段防御模型(早期信号激活→晚期代谢重塑)深化了我们对植物-病原体互作的理解,并为精准育种提供了潜在靶点。(1) 烟草对丁香假单胞菌侵染表现出双相免疫反应。(2) 阶段性转录组重编程协调水杨酸/茉莉酸/生长素信号传导与免疫激活。(3) 次生代谢物生物合成与转录组重编程协同作用。(4) 烟草响应丁香假单胞菌侵染的转录调控网络解析。(5) WRKY6/WRKY23通过调控活性氧及代谢通路增强烟草抗性。篇名: Time-series transcriptome and metabolome profiling Uncovers WRKY6 and WRKY23 as critical regulators in tobacco response to Pseudomonas syringae infection期刊: Journal of Advanced ResearchDOI: 10.1016/j.jare.2026.01.036本研究以烟草品种K326为材料,在六叶一心期使用丁香假单胞菌悬液注射接种叶片。采用多组学技术进行分析:于接种后0、12、24、60 h取样并进行转录组测序,利用HISAT2比对、Salmon定量、DESeq2筛选差异表达基因(DEGs),并进行GO与KEGG富集分析;同时,通过广泛靶向代谢组学(UPLC-MS/MS)和靶向激素检测分析代谢物变化,结合PCA、OPLS-DA等方法筛选差异代谢物(DEMs)。此外,利用WGCNA、时序共表达网络(TO-GCN)及关联分析整合转录组与代谢组数据。通过农杆菌介导的亚细胞定位、病毒诱导的基因沉默(VIGS)、CRISPR/Cas9基因编辑及过表达技术验证WRKY6/23基因功能,并通过qRT-PCR、病原菌生物量测定及DAB/NBT染色分析表型。(1)烟草响应丁香假单胞菌侵染的表现型与生理反应分析烟草在遭遇病原菌侵染时,其防御反应如何具体体现在可见表型与内部生理代谢的动态变化上?研究通过系统监测接种P. syringae pv. tabaci后烟草叶片的症状发展及关键激素与氨基酸水平的时序变化,揭示了侵染早期的信号激活模式。病害症状的时序性发展:接种病原菌后,烟草叶片并未立即出现明显症状。直到24小时(hpi),侵染点周围开始出现浅褐色斑点;至60 hpi,这些斑点颜色加深并扩大。对病斑面积的定量分析证实,受损区域随时间推移呈渐进式扩大,直观反映了病害严重度的逐步升级(图1A)。核心激素与代谢物的动态响应:在0至60 hpi期间,多种与防御相关的激素及前体物质含量发生显著变化。具体而言:- 水杨酸(SA)、茉莉酸(JA)和生长素(IAA) 的含量在侵染过程中持续稳定上升,其中SA的积累最为显著,表明SA信号通路在抗病响应中占据主导地位。
- 作为激素生物合成必需前体的L-色氨酸(TRP)和L-苯丙氨酸(Phe) 也呈现类似的持续积累模式。
- 与此不同,脱落酸(ABA) 的含量变化呈现出独特的两相模式:在侵染最初的12 hpi内轻微下降,随后从24 hpi开始逐渐回升(图1B)。
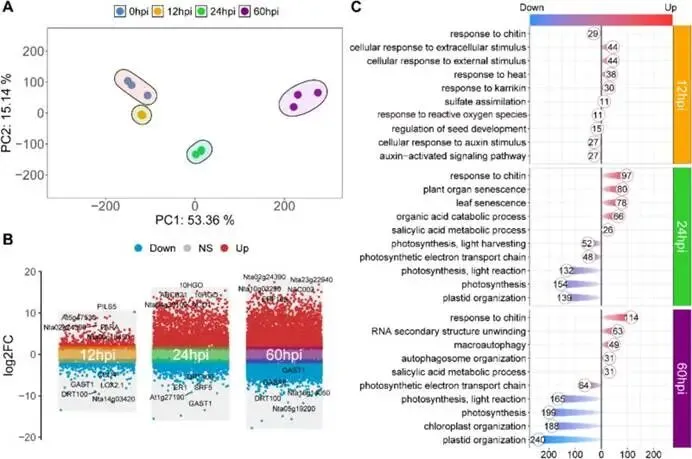
综合结论:烟草对P. syringae的响应伴随着SA、JA等关键防御激素信号的早期、持续激活,以及相关氨基酸前体的同步积累,这为后续的转录与代谢重编程奠定了基础。图1:烟草在P. syringae侵染期间的表现型与生理特征。(A) 烟草叶片在接种P. syringae后0、12、24和60小时(hpi)的表现型变化及病斑面积量化分析。(B)侵染期间烟草体内多种激素的相对丰度,包括水杨酸(SA)、茉莉酸(JA)、生长素(IAA)、脱落酸(ABA)、苯丙氨酸(Phe)和色氨酸(TRP)。数据为三次独立生物学重复的平均值±标准误(N = 3)。面板A中的统计学显著性使用双尾Student’s t检验评估(P < 0.05 (*), P < 0.01 (**), P < 0.001 (***); NS, 不显著)。面板B中的多组比较采用单因素方差分析(ANOVA)及随后的Tukey-Kramer检验;柱状图上方的不同小写字母表示在P < 0.05水平上存在显著差异。(2)响应丁香假单胞菌侵染的烟草时间序列转录组学分析烟草为应对病原菌侵染,其全基因组范围内的基因表达如何随时间发生动态、有序的重编程?研究通过对0、12、24和60 hpi四个时间点的叶片进行RNA-seq测序,结合差异表达基因(DEG)鉴定及功能富集分析,系统解析了防御反应的转录调控时序蓝图。基因表达的全局性动态变化:主成分分析(PCA)显示,不同时间点的样品组内重复性好且组间差异显著(图2A)。与0 hpi对照相比,差异表达基因的数量随侵染进程急剧增加:12 hpi时鉴定到3,031个DEGs,24 hpi时增至10,722个,而到60 hpi时则大幅攀升至20,295个。共有1,624个DEGs在所有时间点均发生差异表达,凸显了侵染对烟草转录组的深远影响(图2B)。已知参与植物激素信号和免疫反应的基因(如SA、JA、IAA通路相关基因及病程相关蛋白基因)在侵染过程中被显著诱导上调,验证了实验数据的可靠性。防御响应的阶段性功能转换:通过GO和KEGG通路富集分析,研究揭示了烟草防御策略随侵染时间发生的根本性转变。- 早期响应(12-24 hpi)——信号感知与应急启动:上调基因主要富集于胁迫响应与激素调节相关功能,如“细胞对细胞外刺激的反应”、“水杨酸代谢过程”。同时,KEGG分析显示氨基酸代谢(缬氨酸、亮氨酸、异亮氨酸降解,色氨酸代谢)和部分次生代谢物生物合成(芪类、二芳基庚烷类、姜辣素)通路高度活跃。这表明植物通过快速调整初级代谢、增强防御激素和特定抗菌化合物的合成来应对初始攻击。
- 后期响应(60 hpi)——稳态维持与能量重分配:上调基因功能转向维持细胞稳态,如“几丁质反应”;同时,整体转录活性受到负调控。从早期开始,与光合作用、光反应和光捕获相关的基因即被显著下调,到60 hpi时抑制加剧。这表明植物在胁迫后期主动降低光合作用能耗,将资源重新分配至防御过程,进入一种节能的自我保护状态(图2C)。
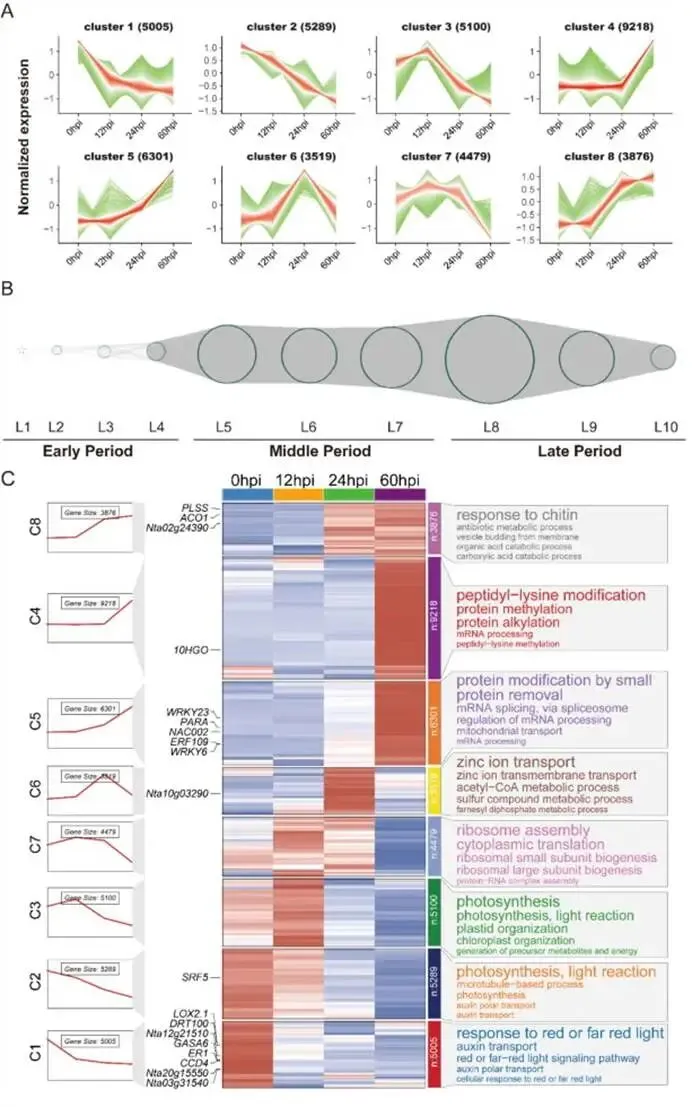
综合结论:烟草的转录组响应呈现出清晰的阶段性:早期侧重于快速信号传导和防御动员,后期则转向代谢重塑和能量节约,构成了一个从“警报”到“适应”的完整防御程序。图2:P. syringae侵染不同时间点的差异表达基因分析。(A) 不同样品基因表达谱的主成分分析(PCA)。(B)火山图展示12 hpi、24 hpi和60 hpi相对于0 hpi的差异表达基因,红点、蓝点和灰点分别代表上调、下调和非显著(NS)基因。(C)12 hpi、24 hpi和60 hpi时间点DEGs的GO富集分析,展示了各时间点上调和下调基因富集的生物过程条目。在复杂的转录重编程背后,是否存在一个有序的基因调控网络来协调不同阶段的防御反应?研究通过Mfuzz聚类和时间顺序基因共表达网络分析,揭示了基因表达的时序模块及其核心调控因子。基因表达模式的聚类分析:Mfuzz方法将所有转录本划分为8个具有不同表达动态的集群(Cluster 1-8)(图3A)。这些集群清晰地反映了侵染进程:Cluster 1-3的基因表达整体下降;Cluster 6和7呈现先升后降的趋势,峰值分别出现在24 hpi和12 hpi;而Cluster 4、5和8的基因表达则在侵染后期持续上调。三级时序调控网络的解析:以bZIP12为起始节点构建的时序基因共表达网络,将DEGs分层到10个时间顺序水平(L1-L10),并进一步归纳为三个时期(图3B):- 早期(L1-L4):基因表达迅速变化,主要涉及光合作用调节和生长素运输,帮助植物快速调整生长-防御平衡。
- 中期(L5-L7):基因表达达到高峰,功能富集于锌离子转运和核糖体组装,关乎能量生产和蛋白质合成,以维持中期感染下的生理功能。
- 晚期(L8-L10):基因持续高表达,功能集中于翻译后蛋白修饰和免疫响应相关通路,标志着防御机制的全面激活。
- 网络分析发现,WRKY、C2H2、ERF和NAC等转录因子家族在各级网络中占据核心位置,同时包括WRKY23、CML19、RPP8L2在内的众多免疫与激素信号相关基因也是网络中的关键节点(图S3C)。
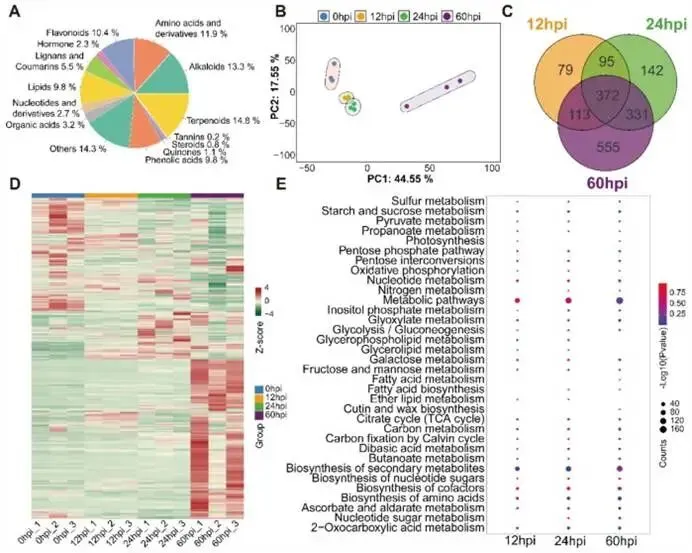
时序模块的功能验证:对8个表达集群的GO/KEGG富集分析结果与上述分期高度吻合(图3C)。例如,早期高表达的Cluster 1-3基因富集于卟啉代谢、淀粉和蔗糖代谢通路;中期Cluster 6、7基因与苯丙氨酸代谢相关;晚期Cluster 4、5、8基因则与谷胱甘肽代谢及缬氨酸/异亮氨酸降解通路有关。这从功能层面证实了时序网络划分的准确性。综合结论:烟草通过一个高度组织化的时序基因共表达网络来协调防御,其中WRKY等核心转录因子家族驱动了从早期信号调整到晚期免疫执行的分阶段转录重编程。图3:响应P. syringae侵染的时序基因共表达网络。(A) 由Mfuzz方法确定的所有差异表达基因聚类结果,显示了在0、12、24和60 hpi时间点的标准化表达模式。(B)预测的烟草响应P. syringae侵染的TO-GCN网络,其中DEGs被组织成10个时间水平(L1–L10),并根据表达动态分为早、中、晚三个时期。(C)时序聚类中基因动态表达变化的热图及其GO富集分析,说明了每个时序聚类相关的代表性基因和富集的生物过程条目。在转录重编程的同时,烟草的代谢组发生了何种全局性重塑?这些代谢变化如何与转录调控相配合,共同构成化学防御体系?研究通过广靶代谢组学和激素代谢组学分析,系统绘制了侵染过程中的代谢图谱。代谢物的整体变化格局:共鉴定到2,768种代谢物,其中生物碱、氨基酸及其衍生物、黄酮类最为丰富(图4A)。主成分分析(PCA)表明,接种后时间差异是导致代谢物组变异的主要因素(图4B)。共有372种差异表达代谢物(DEMs) 在三个侵染后时间点均出现,它们是防御反应中持续起作用的核心代谢物(图4C)。代谢重编程的时序特征:大量DEMs在侵染过程中呈现逐渐增加的模式,尤其是在后期(60 hpi),许多代谢物显著上调(图4D)。KEGG通路富集分析揭示了代谢策略的时序转变:- 侵染早-中期(12-24 hpi):特异性富集的通路包括甘油磷脂代谢、甘油脂代谢和醚脂代谢。这些通路的激活可能与细胞膜重塑、信号分子产生等快速的代谢调整有关,以应对病原菌的初始入侵。
- 侵染中-后期(24-60 hpi):特异性富集的通路转向氮代谢、脂肪酸生物合成、氨基糖和核苷酸糖代谢。这标志着代谢焦点转向防御性化合物的合成,以支持更持久的防御机制(图4E)。
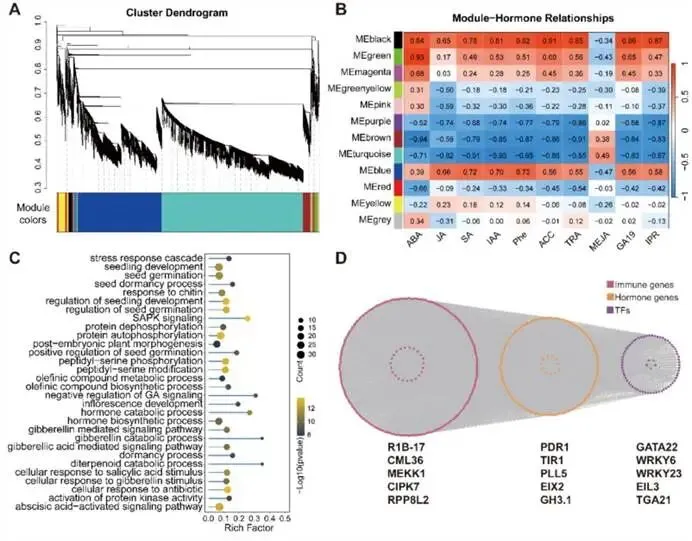
转录组与代谢组的协同分析:整合分析显示,在12 hpi,部分代谢通路(主要是初级和次级代谢过程)在基因表达和代谢物积累水平上同时富集,表明侵染早期即启动了协调的代谢重编程。到24 hpi和60 hpi,两者共同富集的通路数量增加,揭示了在防御中后期,转录调控与代谢输出之间的协同性不断增强(图S4)。综合结论:烟草的代谢防御策略从早期的快速膜代谢和信号调整,过渡到中后期侧重于防御化合物合成的持续代谢重塑,且这一过程与转录重编程紧密耦合。图4:代谢物分类及差异表达代谢物鉴定。(A) 鉴定出的代谢物的分类与组成。(B)基于鉴定代谢物的样本主成分分析(PCA)。(C)P. syringae侵染后差异表达代谢物的维恩图。(D)不同侵染后时间点所有DEMs的热图。(E)不同侵染后时间点DEMs的KEGG富集分析,点大小表示代谢物数量,颜色代表富集显著性。在庞大的基因共表达网络中,哪些模块是连接激素信号与防御反应的核心枢纽?研究通过加权基因共表达网络分析,筛选出与抗病性高度相关的关键基因模块,并解析其调控网络。模块与激素的相关性:WGCNA将全部DEGs划分为12个共表达模块(图5A)。相关性分析发现,黑色、绿色和蓝色模块与SA、JA、IAA、ABA等多种防御激素的含量呈显著正相关;而紫色、棕色和青绿色模块则与激素呈强负相关(图5B)。这提示前三个模块是激素调控网络的核心组成部分。核心模块的功能解析:对黑色、绿色、蓝色模块的深入功能分析揭示了它们各自的特化角色(图5C, S5D, S5E):- 蓝色模块:与赤霉素、水杨酸、脱落酸信号通路以及“蛋白质去磷酸化”过程强烈相关,可能整合多种激素信号,通过蛋白质修饰精确调控下游防御反应。
- 绿色模块:富集于响应光强度、乙烯和细胞分裂素激活的信号通路,可能介导了环境信号(光)与内源激素信号在防御中的交叉对话。
- 黑色模块:核心功能涉及鞘脂代谢过程和活性氧物种响应。鞘脂是细胞膜的重要组成部分和信号分子,其代谢与ROS爆发密切相关,共同构成早期防御屏障。
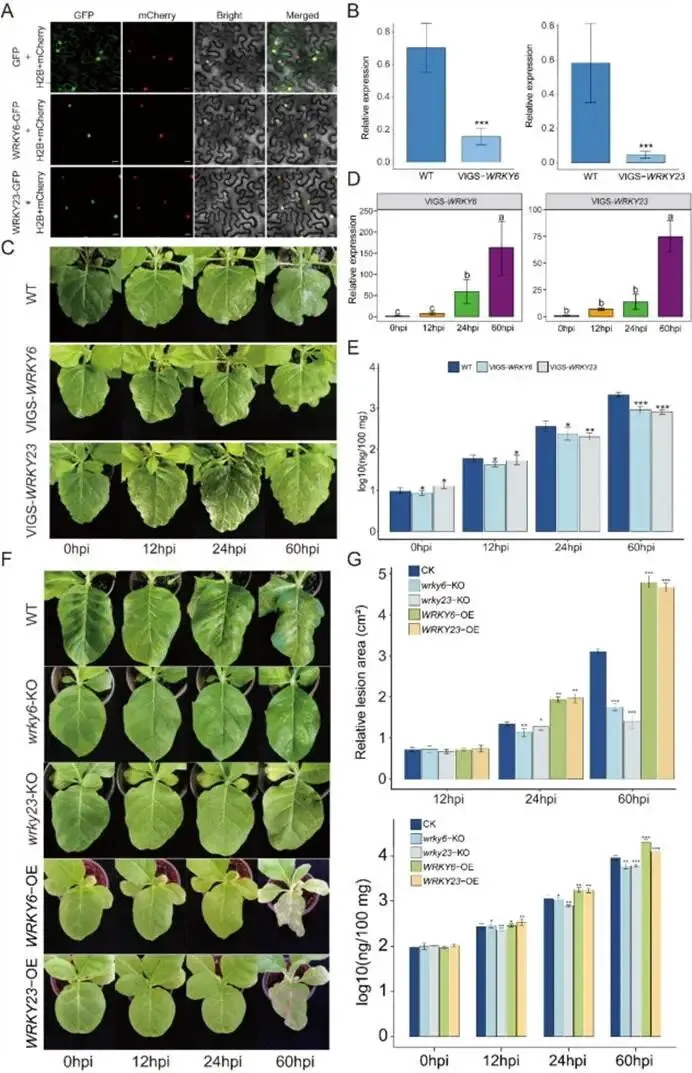
核心模块的调控网络:基因调控网络分析进一步识别出每个模块中的枢纽基因(Hub Genes)。在蓝色模块中,CML36(钙信号)、CIPK7(激酶)、RPP8L2(抗病蛋白)、WRKY23和WRKY6等构成了一个紧密互作的网络核心(图5D)。在黑色和绿色模块中,也分别鉴定出如vps13A、UGT80B1、PRR95、COL9等关键调控因子(图S6B)。这些枢纽基因大多与已知的植物激素和免疫调节相关。综合结论:WGCNA成功鉴定出分别专注于激素信号整合(蓝)、光-激素互作(绿)以及鞘脂-ROS代谢(黑) 的三个核心共表达模块,其中蓝色模块内的WRKY23/WRKY6等枢纽基因成为后续功能验证的优先靶点。图5:烟草P. syringae侵染后的基因模块分析。(A) 显示12个共表达基因模块的聚类树状图。(B)模块与不同激素之间相关性的热图(颜色刻度表示相关性强弱)。(C)蓝色模块基因的GO富集分析,显示显著富集的生物过程条目。(D)蓝色模块中的基因调控网络,突出显示了免疫相关基因、激素相关基因和转录因子。(6)WRKY6与WRKY23作为负调控因子抑制烟草对丁香假单胞菌的抗性基于上述网络分析筛选出的枢纽转录因子WRKY6和WRKY23,它们在抗病中具体扮演何种角色?研究通过亚细胞定位、基因沉默、敲除和过表达等一系列遗传学实验,明确了这两个转录因子的生物学功能。WRKY6与WRKY23的核定位:瞬时表达WRKY6-GFP和WRKY23-GFP融合蛋白在本氏烟草中显示,两者的荧光信号均特异性地定位于细胞核(图6A),符合其作为转录因子的预期。基因沉默与敲除增强抗病性:利用病毒诱导的基因沉默(VIGS)技术构建的VIGS-WRKY6和VIGS-WRKY23植株,其靶基因表达量显著降低(图6B)。接种病原菌后,与野生型(WT)相比,沉默植株的病斑面积更小,叶片内的病原菌DNA含量显著降低(图6C, E)。利用 CRISPR-Cas9 技术创制的wrky6-KO和wrky23-KO纯合突变体进一步证实了这一结果:突变体植株的病斑扩展和细菌载量均显著低于WT(图6F, G)。基因过表达增加感病性:与之相反,WRKY6-OE和WRKY23-OE过表达株系在接种后表现出更严重的黄化症状和更大的病斑面积,叶片内病原菌积累量也显著高于WT(图6F, G)。综合结论:遗传学证据一致表明,WRKY6和WRKY23是烟草对P. syringae抗性的负调控因子。抑制其功能能增强植物抗性,而过量表达则导致感病性增加。图6:WRKY6与WRKY23介导烟草中核定位的、通过ROS信号对抗P. syringae的防御响应。(A)WRKY6和WRKY23的亚细胞定位。本氏烟草叶片瞬时转化融合构建体(WRKY6-GFP和WRKY23-GFP)及核标记H2B-mCherry。(B)WT与VIGS-WRKY6/VIGS-WRKY23沉默植株中WRKY6和WRKY23的相对表达水平。(C)P. syringae侵染后WT与VIGS-WRKY6/VIGS-WRKY23沉默植株的表现型分析。(D)VIGS-WRKY6/VIGS-WRKY23沉默植株在P. syringae侵染期间WRKY6和WRKY23的时序表达模式。(E)使用物种特异性引物对定量受侵染叶片中的P. syringae DNA。(F)WT、wrky6-KO、wrky23-KO、WRKY6-OE和WRKY23-OE植株在接种P. syringae后0、12、24和60 hpi的表现型分析。(G)WT、敲除突变体和过表达株系在0、12、24和60 hpi的相对病斑面积和P. syringae DNA水平量化。数据为三次独立生物学重复的平均值±标准误(N = 3)。面板B和E中的统计学显著性使用双尾Student’s t检验评估(P < 0.05 (*), P < 0.01 (**), P < 0.001 (***); NS, 不显著)。面板D中的多组比较采用单因素方差分析(ANOVA)及随后的Tukey-Kramer检验;柱状图上方的不同小写字母表示在P < 0.05水平上存在显著差异。(7)WRKY6与WRKY23通过调控ROS及苯丙烷代谢调节烟草抗性WRKY6与WRKY23作为转录因子,其负调控抗性的分子机制是什么?它们是否通过影响下游的活性氧爆发或特定代谢途径来发挥作用?研究通过组织化学染色、代谢物关联分析及分支代谢突变体验证,深入阐释了其下游作用通路。调控活性氧积累:通过DAB(检测H2O2)和NBT(检测O2•–)染色发现,在接种后48 hpi,VIGS-WRKY6和VIGS-WRKY23沉默植株叶片中ROS的积累水平显著高于WT植株(图7A)。增强的ROS爆发有助于限制病原菌扩展,这直接解释了沉默植株和敲除突变体抗性增强的表型。关联代谢通路与关键代谢物鉴定:相关性分析显示,与WRKY6/23表达最显著相关的代谢物主要属于氨基酸及其衍生物类别。KEGG富集分析指出,WRKY6/23可能特别调控苯丙氨酸代谢等通路(图7B)。其中,苯丙烷代谢通路中的L-苯丙氨酸及其下游防御性化合物黄嘌呤核苷(xanthosine),与WRKY6/23的表达水平呈极强的正相关(图7B)。转录组数据也证实,L-苯丙氨酸和黄嘌呤核苷分支上的众多基因受侵染诱导且与WRKY6/23表达相关。分支代谢的遗传与药理学验证:为了明确这两个代谢分支的功能,研究创制了相应的突变体并进行外源处理:- L-苯丙氨酸分支:敲除该分支关键基因MAGL3的MAGL3-ko突变体,在接种后表现出病斑增大、细菌载量升高,即更感病。相反,外源施加L-苯丙氨酸能减轻WT叶片的病害症状并降低细菌数量(图7C-E)。
- 黄嘌呤核苷分支:敲除该分支关键基因Mik的Mik-ko突变体,则显示出病斑减小、细菌载量降低,即更抗病。而外源施加黄嘌呤核苷会加重WT叶片的病害(图7C-E)。
综合结论:WRKY6和WRKY23通过负调控ROS的爆发,并精细调节苯丙烷代谢分支的流向来影响抗性。它们促进感病性分支产物(黄嘌呤核苷)的积累,同时可能抑制抗病性分支前体(L-苯丙氨酸)的利用或转化,从而将转录调控与代谢防御网络直接联系起来。图7:WRKY6与WRKY23敲除突变体对P. syringae的感病性及时序基因表达谱。(A) 在接种P. syringae后48 hpi,WT和VIGS-WRKY6/VIGS-WRKY23沉默植株离体叶片中H2O2(DAB染色)和O2–(NBT染色)的组织化学检测。(B)与WRKY6和WRKY23相关代谢物的KEGG分析。(C)野生型(CK)、MAGL3敲除、Mik敲除及代谢物预处理(L-苯丙氨酸+和黄嘌呤核苷+)烟草叶片在接种P. syringae后0、12、24和60 hpi的代表性表型。(D)野生型、敲除突变体及代谢物预处理植株叶片在12、24和60 hpi的相对病斑面积量化。(E)各基因型和处理植株受侵染叶片在0、12、24和60 hpi的细菌数量量化。数据为三次独立生物学重复的平均值±标准误(N = 3)。所有统计学比较均使用双尾Student’s t检验;显著性表示为P < 0.05 (*), P < 0.01 (**), P < 0.001 (***); NS, 不显著。本研究系统阐明了烟草在丁香假单胞菌侵染过程中激活的多层次防御机制。转录重编程的时序动态揭示了一种双相响应策略:在病原菌入侵后,植物通过快速激活胁迫响应通路及激素信号(水杨酸、茉莉酸和生长素)实现防御优先化;而关键次生代谢物(尤其是生物碱和类黄酮)生物合成通路的持续诱导,则凸显了其在化学防御中的核心作用。WRKY、ERF和NAC等多个转录因子家族在协调阶段特异性调控网络中的核心功能,进一步证实了它们作为植物免疫关键节点的进化保守性。对WRKY6与WRKY23的功能验证确证了二者作为抗性负调控因子的作用,其基因沉默能增强活性氧爆发及L-苯丙氨酸、黄嘌呤核苷等关键下游代谢物的积累。沉默株系中病原菌生物量的显著降低不仅证实了其生物学重要性,更确立了这两个转录因子作为精准育种潜在靶点的价值。然而,野生型植株中未出现完全抗性,表明存在额外的调控层级,可能涉及与未知转录因子家族的互作或转录后修饰机制。未来研究应优先聚焦于这些调控网络的时空解析,并在重要经济作物中验证其保守性。本工作为理解植物-病原互作提供了新见解,并为通过分子育种策略培育新一代抗病烟草品种建立了坚实框架。


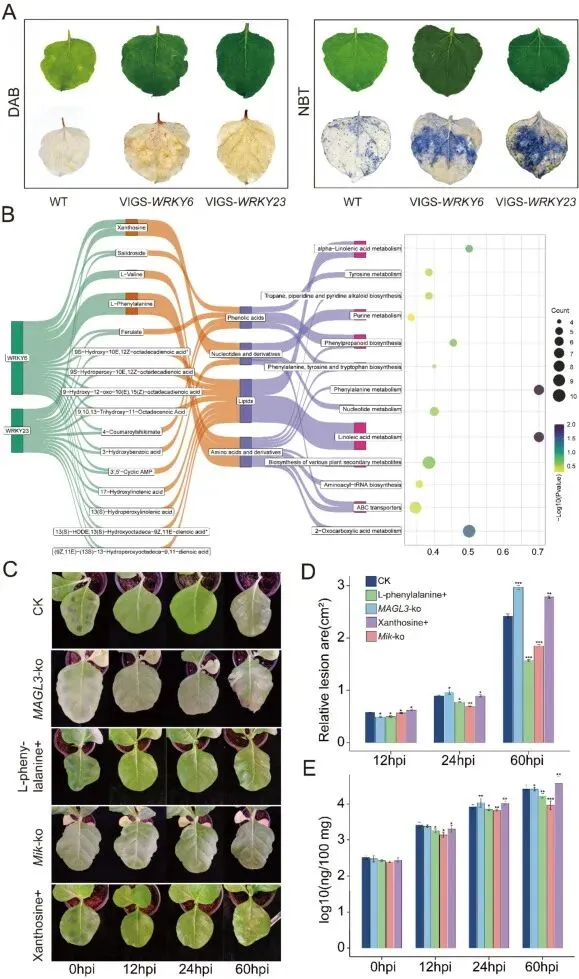





 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
