2026年2月5日,郑州大学生命科学学院于政权团队联合中国科学院深圳先进技术研究院王枫、中国农业大学吕聪以及加州大学尔湾分校Maksim V. Plikus等四家单位,在干细胞领域顶级期刊《Cell Stem Cell》在线发表了题为《Sleep disturbance triggers aberrant activation of vagus circuitry and induces intestinal stem cell dysfunction》的研究长文。
该研究首次系统阐明了“睡眠紊乱—脑—肠轴—干细胞”这一全新病理链路,为理解睡眠障碍如何远程、精准地削弱肠道自我更新能力提供了分子细胞层面的全景图,也为防治炎症性肠病、代谢性胃肠并发症等提出了可干预的靶点。以下从研究背景、技术路线、核心发现、机制解析、意义与展望五个维度进行深度解读。
一、研究背景:睡眠与肠道为何“同病相怜”
临床上长期观察到,夜班工人、失眠患者及跨时区旅行者出现胃肠道症状的概率显著升高,但“睡不好”究竟如何跨越血脑与肠黏膜两大屏障,最终削弱肠道上皮更新,始终是悬而未决的问题。过去十年,脑-肠轴研究主要聚焦“微生物—免疫”或“神经—免疫”交互,而干细胞作为肠道稳态的“建筑工人”,其是否及如何接收中枢睡眠信号却鲜有报道。该团队前期发现,慢性睡眠剥夺可诱发果蝇肠干细胞(ISC)过度增殖却修复障碍(Zhou et al., Cell Prolif 2023)[3],提示睡眠-干细胞轴可能保守存在。于是,作者提出假说:睡眠中断通过迷走神经介导的脑-肠回路,远程诱导肠道干细胞氧化应激与功能耗竭,从而破坏上皮屏障。
二、技术路线:四单位互补的“跨界”组合拳
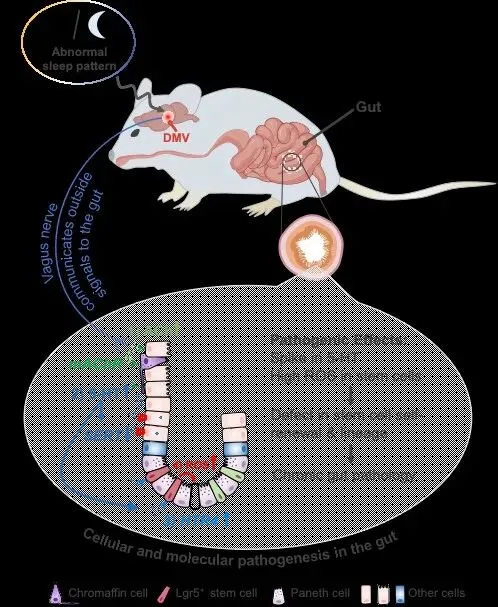
1. 模型构建:采用“环境+遗传”双策略。① 睡眠干扰模型:小鼠置于自动睡眠碎片化平台,每日睡眠剥夺70%以上,持续4周;② 迷走神经特异性干预:利用Vgat-Cre、Chat-Cre、Phox2b-Cre等小鼠,交叉AAV-DIO-hM4Di或hM3Dq,实现迷走神经节神经元化学遗传学抑制/激活;③ 干细胞命运追踪:Lgr5-GFP、Axin2-CreERT2、Bmi1-CreERT2报告小鼠,结合EdU脉冲标记、类器官单克隆谱系追踪,量化干细胞自我更新与分化平衡。
2. 多组学解析:① 空间转录组(Visium)定位睡眠剥夺后十二指肠隐窝-绒毛轴基因表达差异;② 单细胞核RNA-seq(snRNA-seq)解析迷走神经节、孤束核及肠神经丛细胞亚群转录改变;③ 靶向代谢组检测肠黏膜ROS、GSH/GSSG、NADPH/NADP+及脂质过氧化终产物。
3. 干预验证:① 迷走神经切断(subdiaphragmatic vagotomy);② 抗氧化剂NAC或线粒体靶向MitoQ灌胃;③ 肠上皮特异性FoxO3或Nrf2敲除,检验氧化应激在干细胞功能缺损中的必要性;④ 类器官共培养:将睡眠剥夺小鼠血清或迷走神经节条件培养基作用于健康隐窝类器官,观察“远程伤害”是否足以直接诱发干细胞缺陷。
三、核心发现:一条从“睡眠中枢”到“隐窝干细胞”的五级信号链
1. 睡眠碎片化导致迷走神经背核(DMV)异常兴奋:c-Fos+神经元比例升高2.3倍,电生理记录显示其向胃肠道的传出放电频率增加近50%。
2. DMV异常信号经迷走神经外周支传递至肠道:在体光纤钙成像显示,睡眠剥夺小鼠胃肠道外周迷走轴突Ca2+瞬变幅度显著增强;化学遗传学抑制DMV可阻断该现象。
3. 肠道接收到“应激指令”后,首先激活间质细胞(telocytes)释放乙酰胆碱(ACh)与NO,引发局部血管舒张及免疫细胞浸润;单细胞数据发现,Col18a1+间质细胞是早期响应迷走信号的“第一哨兵”。
4. 干细胞微环境氧化应激水平升高:睡眠剥夺组隐窝内8-oxo-dG与脂质过氧化终产物MDA分别升高1.8倍和2.1倍;线粒体形态由管状变为碎片状,膜电位下降。
5. 隐窝干细胞功能受损:EdU标记显示,睡眠剥夺使Lgr5+干细胞静止期(G0)比例下降34%,但增殖后无法有效分化为分泌型细胞;类器官形成效率降低42%,传代潜能下降3代;FoxO3核转位减少,抗氧化基因Sod2、Gclc表达下调。
四、机制解析:迷走神经-氧化应激轴是“罪魁祸首”
1. 迷走神经的必要性:膈下迷走神经切断可完全阻断睡眠剥夺诱导的干细胞增殖-分化失衡及上皮通透性增加,证明迷走通路是“脑-肠”传递的必需环节。
2. 氧化应激的充分性:睡眠剥夺小鼠口服NAC或MitoQ,可恢复隐窝干细胞线粒体膜电位,逆转类器官形成缺陷;而肠上皮特异性缺失FoxO3后,抗氧化剂的保护效应消失,提示FoxO3是迷走信号-氧化应激-干细胞轴中的关键转录节点。
3. 人源化证据:团队与加州尔湾分校合作,采集20例慢性失眠患者及20名匹配健康志愿者的十二指肠活检标本,发现失眠组隐窝8-oxo-dG水平与PSQI评分呈正相关,且Lgr5表达降低;离体培养显示,患者血清可使健康结肠类器官出芽减少,而迷走神经阻断剂阿托品或抗氧化剂可拮抗该效应,为机制的人体相关性提供了直接证据。
五、意义与展望:把“睡眠医学”推向“干细胞保护”前沿
1. 理论层面:首次绘制了“睡眠中枢-迷走神经-间质细胞-干细胞微环境-氧化应激”五级因果链,填补了脑-肠轴研究在再生医学层面的空白。
2. 临床层面:为炎症性肠病、化疗相关肠黏膜炎、糖尿病胃肠病变等“高氧化应激+上皮修复障碍”疾病提供了新的病因学视角,提示改善睡眠或靶向迷走神经-抗氧化通路可作为辅助疗法。
3. 干预策略:① 精准干预:化学遗传学或超声迷走神经调控,已在癫痫、抑郁领域开展临床实验,未来可拓展至胃肠保护;② 药物重定位:FDA已批准的迷走神经拮抗剂(如阿托品、右美托咪定)及线粒体靶向抗氧化剂(MitoQ、SS-31)可快速进入 repurposing 研究;③ 生活方式:研究提示“深度睡眠>6 h”即可显著降低隐窝氧化应激,公共卫生指南可将“保护肠道干细胞”纳入睡眠推荐的新论据。
4. 未来方向:① 迷走神经不同亚型(胆碱能、肽能、嘌呤能)在睡眠-肠轴中的功能异质性;② 肠道菌群-迷走-脑逆向信号是否放大睡眠障碍的恶性循环;③ 是否可利用迷走神经刺激(VNS)主动增强干细胞功能,用于放射性肠炎或短肠综合征的再生治疗。
综上,该研究以干细胞为核心切入点,跨越神经科学、消化生理与再生医学的学科壁垒,为解释“睡不好就拉肚子”这一日常现象提供了严谨的科学证据,更为睡眠医学与胃肠病学的交叉研究树立了新标杆。随着脑-肠-干细胞轴研究的深入,我们或许能在不久的将来看到“睡眠处方”成为肠道疾病标准治疗的一部分。
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
