IF42.5!苏州大学&郑州大学用“国自然”荣登《Cell》顶刊!揭示AARS1乳酰化p53促癌机制!
- 2026-06-13 14:52:45





乳酸是糖酵解的代谢产物,近年发现其可通过组蛋白乳酸化调控基因表达,但细胞内乳酸如何被感知并转化为蛋白质乳酸化的机制尚不明确。苏州大学团队通过全基因组CRISPR筛选发现,丙氨酰-tRNA合成酶AARS1是细胞内乳酸传感器和乳酸转移酶,直接结合乳酸并通过ATP依赖方式催化全局赖氨酸乳酸化。
AARS1介导p53的K120和K139位点乳酸化,抑制p53的液-液相分离、DNA结合和转录活性,促进肿瘤发生。β-丙氨酸可竞争性抑制乳酸与AARS1结合,阻断p53乳酸化并抑制肿瘤生长。该研究揭示了代谢物乳酸直接调控蛋白质功能的新范式。
📢如果你也想用“国自然”发文的话,关注西瓜生信,联系我们带你进一步了解~


文章标题:丙氨酰-tRNA合成酶AARS1是乳酸传感器和乳酸转移酶,通过乳酸化p53促进肿瘤发生
发表期刊:Cell
发表时间:2024年4月22日
影响因子:IF42.5/Q1

研究背景

瓦博格效应是癌症的标志性代谢特征,肿瘤细胞即使在有氧条件下也依赖糖酵解产生大量乳酸。传统认为乳酸是代谢废物,但近年发现乳酸可通过组蛋白乳酸化修饰调控基因表达。抑制p53的液-液相分离、DNA结合和转录活性,从而促进肿瘤发生。β-丙氨酸可竞争性抑制该过程,为癌症治疗提供新策略。

研究方法

本研究通过TCGA数据库分析发现乳酸与p53信号通路负相关,体内外实验证实乳酸抑制p53活性并促进p53乳酸化。全基因组CRISPR筛选鉴定出AARS1为介导p53乳酸化的关键酶。采用MST、CD、EMSA、FRAP、LC-MS/MS等方法系统阐明AARS1催化乳酸-AMP形成及转移至赖氨酸的生化机制。

你也想挑战前沿生信研究方向?你也想拥有同款高分数据库,却苦于不知从何入手?别再观望了,赶紧联系西瓜生信团队!我们提供全方位专业支持,从数据分析到思路设计,助你轻松搞定科研难题!

肿瘤来源的乳酸是p53的天然抑制物,通过乳酸化抑制p53
TCGA乳腺癌数据分析显示,高乳酸信号与p53信号通路评分显著负相关。在MMTV-PyMT乳腺癌小鼠模型中,腹腔注射乳酸钠加速肿瘤发生并抑制p53活性。LDHA条件性敲除延迟肿瘤形成、降低肿瘤乳酸含量并增加p53活性(图1)。
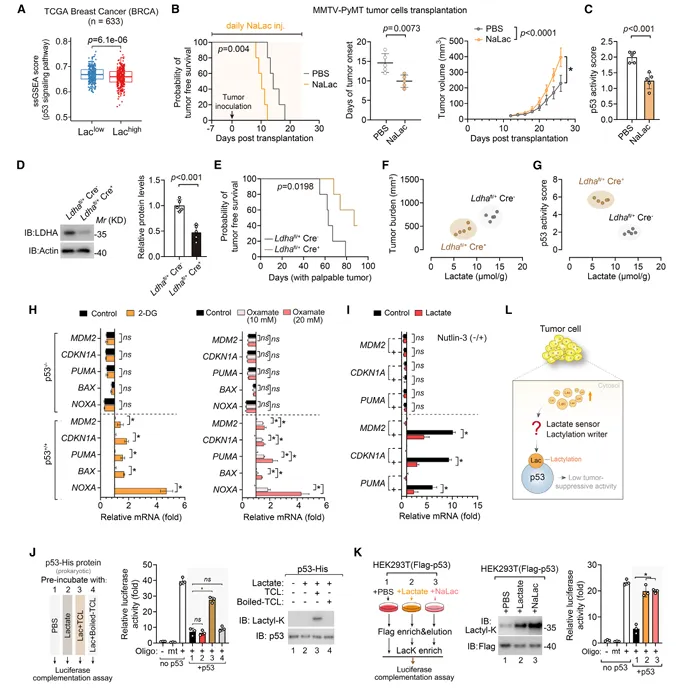
体外实验中,乳酸或乳酸钠处理可诱导p53乳酸化,而糖酵解抑制剂2-DG或LDHA抑制剂oxamate减少p53乳酸化并增强p53靶基因表达。利用细胞裂解液而非纯乳酸直接处理p53才能降低其活性,且降低程度与乳酸化水平相关,提示存在催化乳酸化的酶。
全基因组CRISPR筛选鉴定AARS1为全局赖氨酸乳酸化的介导者
构建p53-GFP报告基因的HeLa细胞,经乳酸和Nutlin-3处理后分选GFP高表达细胞进行CRISPR筛选,AARS1是最显著的候选基因。AARS1敲低增强p53报告基因活性和靶基因表达,抑制细胞增殖和集落形成,并阻断乳酸诱导的促肿瘤效应(图2)。
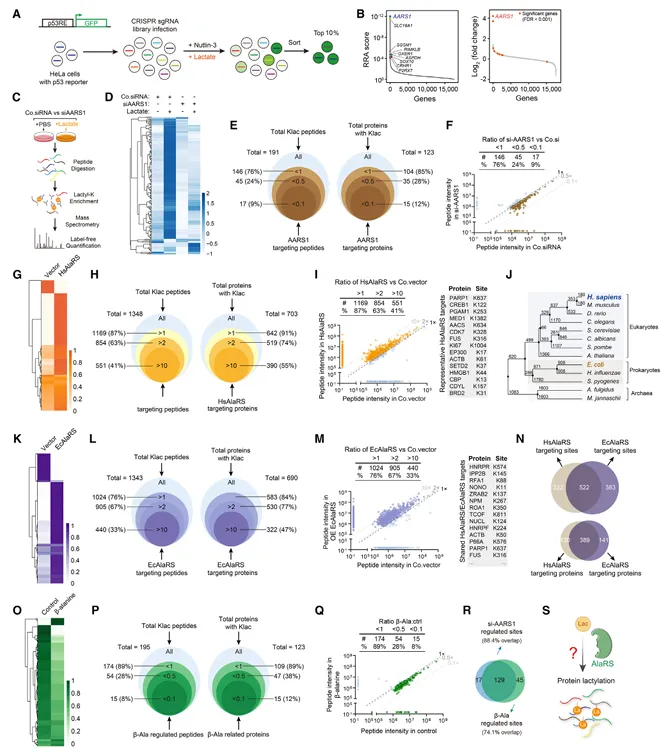
图2.全基因组CRISPR筛选鉴定AARS1为全局赖氨酸乳酸化的介导者
蛋白质组学分析显示,AARS1敲低使约80%的乳酸化肽段强度下降,而AARS1过表达使90%的乳酸化肽段强度升高。大肠杆菌EcAlaRS同样具有催化真核细胞乳酸化的能力,且与AARS1的靶点高度重叠。β-丙氨酸可有效抑制全局乳酸化。
AARS1直接结合乳酸并以ATP依赖方式催化赖氨酸乳酸化
MST分析显示EcAlaRS和HsAlaRS与乳酸的Kd分别约为13 μM和35 μM,β-丙氨酸亲和力更高并可竞争性抑制乳酸结合。分子对接鉴定出保守的乳酸结合残基,突变这些残基完全消除乳酸结合和乳酸化催化活性(图3)。
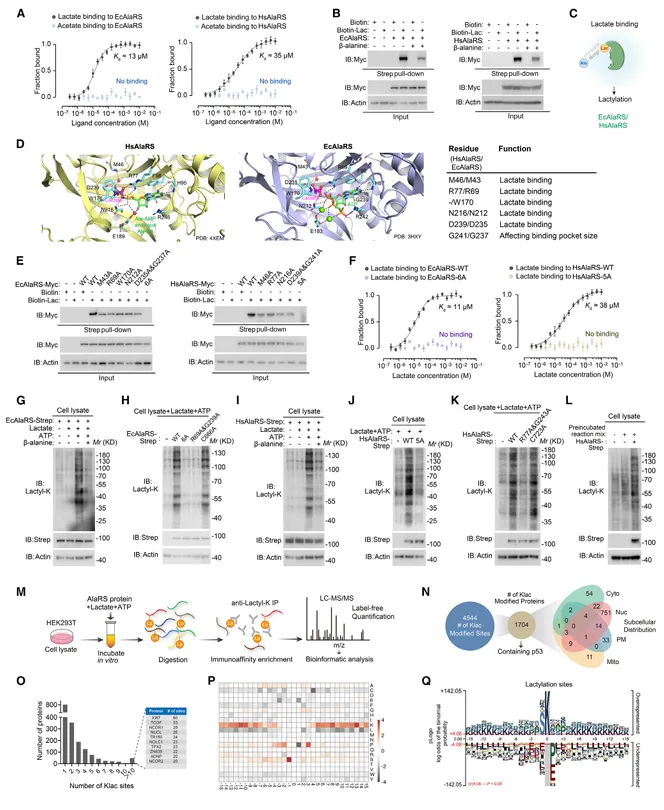
图3.AARS1直接结合乳酸并以ATP依赖方式催化赖氨酸乳酸化
体外反应需要同时存在乳酸、ATP和AARS1,且ATP结合突变体同样失活。通过去除AARS1后无法催化乳酸化,证实AARS1直接转移乳酸基团而非产生可扩散中间体。质谱鉴定出EcAlaRS催化的4,544个乳酸化位点,覆盖1,704个蛋白,乳酸化位点周围氨基酸序列具有偏好性。
AARS1通过ATP依赖的乳酸-AMP中间体将乳酸共价转移至赖氨酸
蛋白质组学显示AARS1是p53的结合蛋白,Co-IP证实两者直接相互作用,Kd约为39 μM和21 μM。在细胞和体外,AARS1-WT而非乳酸或ATP结合突变体可催化p53的K120和K139位点乳酸化,并被β-丙氨酸抑制(图4)。
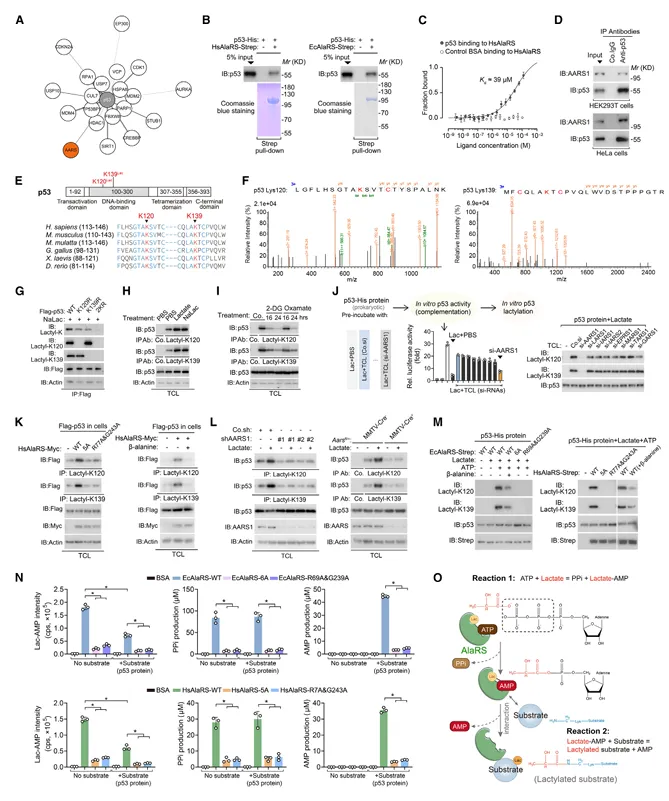
图4.AARS1通过ATP依赖的乳酸-AMP中间体催化赖氨酸乳酸化
AARS1催化乳酸和ATP生成乳酸-AMP并释放焦磷酸,加入底物p53后乳酸-AMP被消耗并产生AMP。AARS1对乳酸的催化效率约为丙氨酸的1/2000,但细胞内乳酸浓度远高于丙氨酸,足以支持乳酸化发生。
p53位点特异性乳酸化削弱其DNA结合和液-液相分离,降低肿瘤抑制功能
p53的K120和K139位于DNA结合域。通过遗传密码扩展技术构建位点特异性乳酸化p53蛋白。EMSA和MST显示,p53Dual-Lac与p53RE-DNA的亲和力降低约1000倍,p53K120-Lac降低约100倍,p53K139-Lac降低约10倍(图5)。
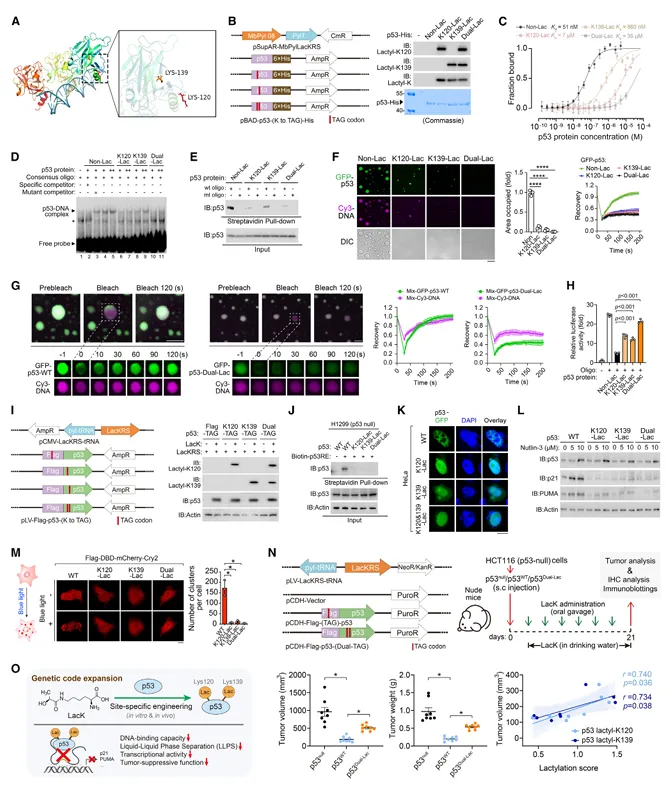
图5.p53位点特异性乳酸化削弱其DNA结合和LLPS
p53Non-Lac与p53RE-DNA可形成液滴,而乳酸化变体形成液滴能力显著减弱。在p53-/-细胞中,表达乳酸化p53的细胞无法结合p53RE-DNA、无法形成核内凝聚体、转录活性降低,且在小鼠体内更具致瘤性。肿瘤中p53乳酸化水平与肿瘤体积正相关。
模拟乳酸化的K120/K139癌症相关突变体表现出LLPS和肿瘤抑制能力降低
分析TP53数据库和COSMIC数据库发现,p53的K120和K139存在多种癌症相关突变突变模拟了乳酸化对电荷的改变(图6)。
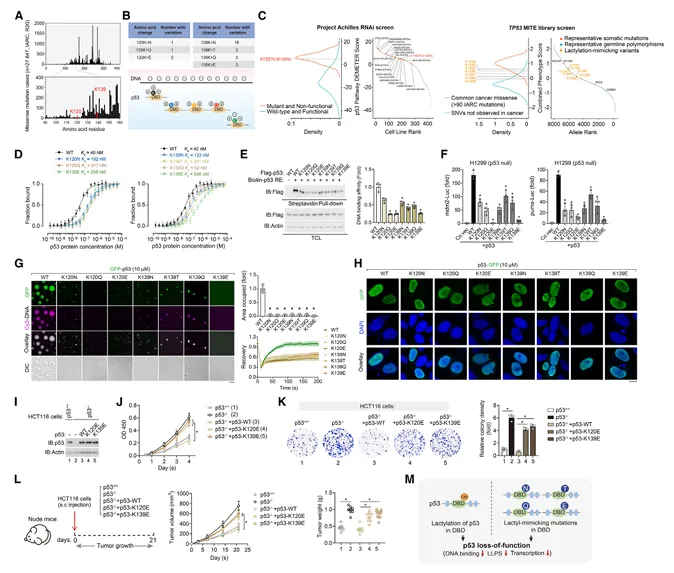
图6.疾病相关的K120/K139模拟乳酸化突变体表现出LLPS和肿瘤抑制能力降低
功能实验显示,K-to-E突变体与p53RE-DNA结合力丧失最严重,转录活性几乎完全消失,LLPS能力受损,在p53-/-细胞中无法抑制增殖、集落形成和异种移植瘤生长。此外,乳酸化与乙酰化在相同位点相互排斥:乳酸化p53不能被乙酰化,乙酰化p53也不能被乳酸化,而乙酰化与乳酸化功能相反。

本研究首次揭示丙氨酰-tRNA合成酶AARS1是细胞内乳酸传感器和乳酸转移酶,通过ATP依赖的乳酸-AMP中间体催化全局赖氨酸乳酸化,并特异性乳酸化p53的K120和K139位点。乳酸化通过中和赖氨酸正电荷,破坏p53 DNA结合域与响应元件的结合,抑制p53的液-液相分离、转录活性和肿瘤抑制功能。
由于AARS1在多种癌症中高表达且与不良预后相关,靶向AARS1-乳酸化轴可能成为广谱抗癌新策略,但需注意AARS1同时具有翻译功能,需选择性干预其乳酸转移酶活性。
📢如果你也想用“国自然”进行发文,思路复现,那就快来联系我们吧~

有需要的朋友联系~

IF30.9!南京大学&南京医科大学联手用“国自然”荣登《Cell》子刊!竟发现父亲运动可通过精子miRNA提升后代耐力!

随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 产教融合聚智赋能 协同共建汽车强省——郑州航空港区数智制造产业协会协办河南省新能源与智能网联汽车专业集群建设研讨会圆满落幕
- 科室简介丨郑州大学第五附属医院精神心理科
- 郑州户外旅游群活动总汇:2026年4月14号更新
- 郑州市管城回族区二里岗社区卫生服务中心岗位招聘公告
- 比亚迪郑州直招,超多岗位(校招+社招+普工招聘)
- 郑州航空港区银河社区卫生服务中心招聘公告!
- 郑州进入流感高发季!严重可致命!甲型H1N1为主,出现这几个症状快就医...
- 每周资讯精选 | 郑州入选!国务院批复部分地区要素市场化配置改革试点;全国首个省级PPP存量项目建设运营方案发布
- 郑州灵活就业医保可以转为居民医保吗?
- 因雨水管线施工,郑州一道路临时导改为单向通道!
