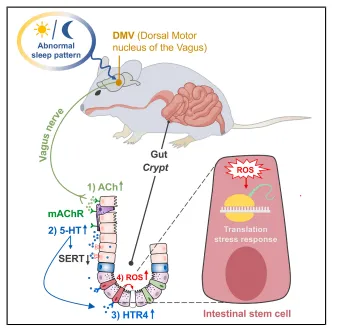
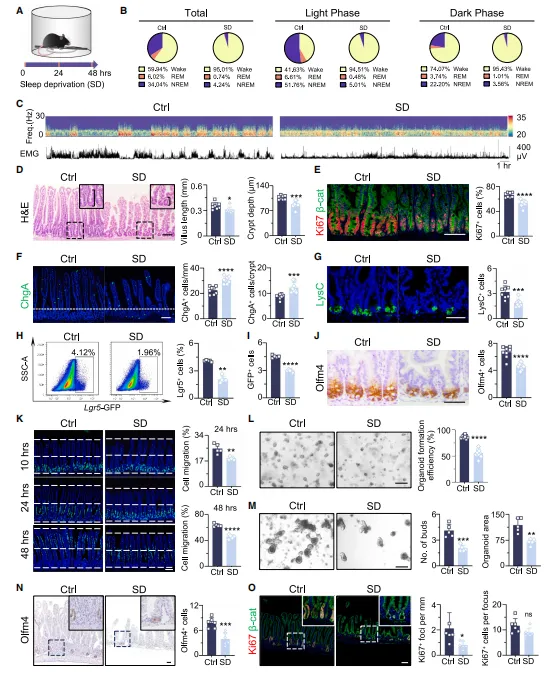
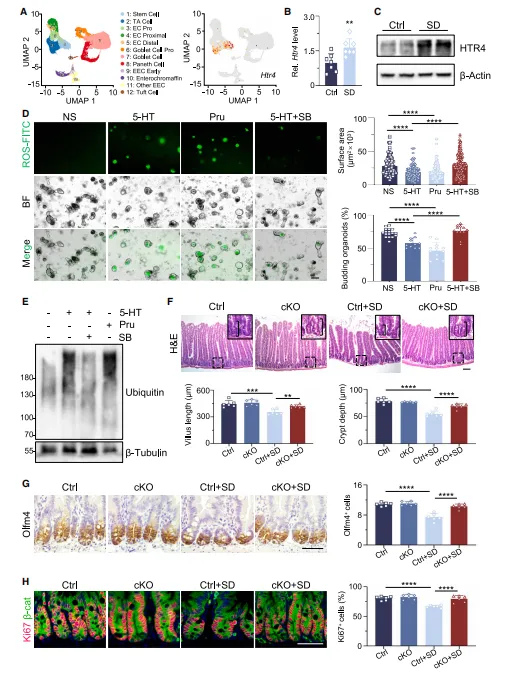
小鼠连续两天睡眠不足后,肠道里的绒毛和隐窝都明显变短了,肠道干细胞、潘氏细胞数量也随之减少。细胞标记追踪发现,肠道细胞向上迁移的速度变慢,意味着肠道自身的更新修复能力大打折扣。后续体外培养实验也印证了这一点:缺觉小鼠的肠道隐窝,很难再培育出正常的肠道类器官(图1)。
图1.短期SD显著损害ISC功能并破坏肠上皮细胞更新
复现思路:
想验证一个处理是否影响肠道干细胞,别只看HE染色。建议加上Lgr5或Olfm4的免疫组化,再用BrdU脉冲示踪看细胞迁移。这三步做下来,结论就比较扎实了。
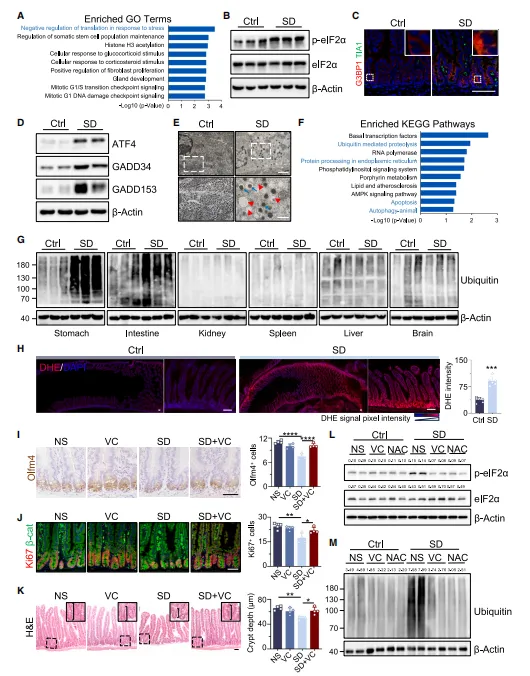
蛋白检测结果发现,睡眠不足会让肠道隐窝启动翻译抑制的应激反应。相关应激信号、应激小体明显变多,多种应激相关蛋白水平也随之升高;同时细胞内有害自由基增多,蛋白异常降解也加剧了。而补充维生素 C 或 NAC 后,这些异常情况都能得到改善(图2)。
图2.短期SD通过激活翻译应激反应损害ISCs功能
复现思路:
拿到处理组和对照组的组织后,不妨先做个非靶向蛋白组学,看看哪些通路富集。缺觉这种全身性干预,直接猜分子容易跑偏,让数据告诉你该看什么~
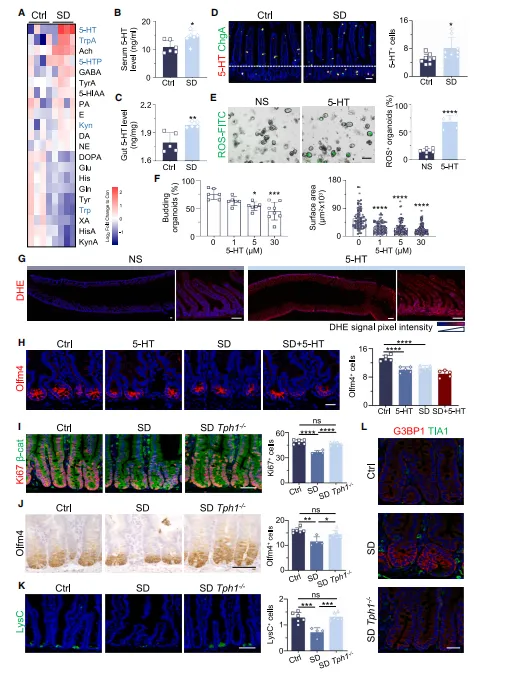
长期睡眠不足的小鼠,肠道和血液里的5 - 羟色胺含量明显变高,肠道分泌这类物质的 EC 细胞数量也随之增多。
实验发现,给肠道类器官外加 5-HT 后,细胞内活性氧会增多,肠道正常生长也受到抑制;往小鼠体内注射 5-HT,还能直接模拟出睡眠不足引发的肠道异常状态。
而敲除合成 5-HT 关键基因 Tph1 的小鼠,即便长期缺觉,肠道也基本不会出现损伤(图3)。
图3.急性SD可破坏5-HT稳态并诱导肠道氧化应激
复现思路:
如果怀疑某个生理状态改变会影响肠道,可以先把神经递质扫一遍,用LC-MS/MS做靶向代谢组学。
单细胞检测发现,Htr4 基因在肠道干细胞中活跃度很高。睡眠不足时,肠道里对应的 HTR4 蛋白含量也会明显增加。实验中,用 HTR4 激活剂处理肠道类器官,会和 5 - 羟色胺一样抑制肠道正常生长;而使用抑制剂就能逆转这种不良影响。针对肠道专门敲除 Htr4 基因后,小鼠就算熬夜缺觉,肠道干细胞受到的损伤也大幅减轻了(图4)。
图4.间充质干细胞通过其受体HTR4响应5-羟色胺
复现思路:
找到关键配体后,下一步自然是找受体。先用公共单细胞数据看受体表达谱,锁定候选靶点,再用类器官体系做配体和拮抗剂的交叉验证。
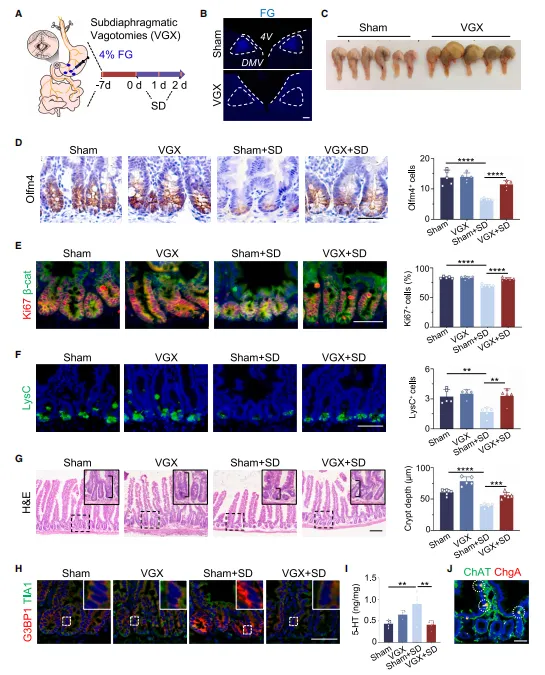
给小鼠切断双侧迷走神经后,即便再让它睡眠不足,其肠道干细胞数量也能基本恢复正常,肠道隐窝深度回归常态,体内 5-HT 水平也不再异常升高。染色结果显示,缺觉会刺激大脑多个区域活跃,而仅 DMV 脑区同时被示踪病毒标记,证明这个脑区和肠道之间存在直接的神经通路(图5)。
复现思路:
想证明某个脑区到外周器官的功能连接,先用逆行病毒看解剖上的连接,再用手术切断神经看功能是否依赖这条通路。
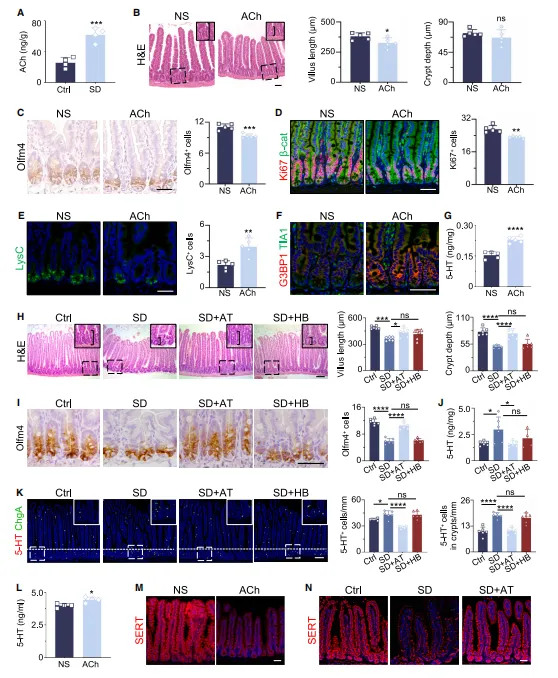
睡眠不足的小鼠,肠道内乙酰胆碱含量会增多。给正常小鼠直接注射乙酰胆碱后,它体内的 5-HT 也会随之升高,肠道状态和缺觉的小鼠表现完全一致。使用阿托品处理后,就能阻止 5-HT 异常升高,保护肠道健康;但烟碱受体阻断剂没有这类效果(图6)。
复现思路:
找到神经通路后,下一步就是鉴定下游的神经递质和受体。体内拮抗剂筛选是比较经济高效的做法
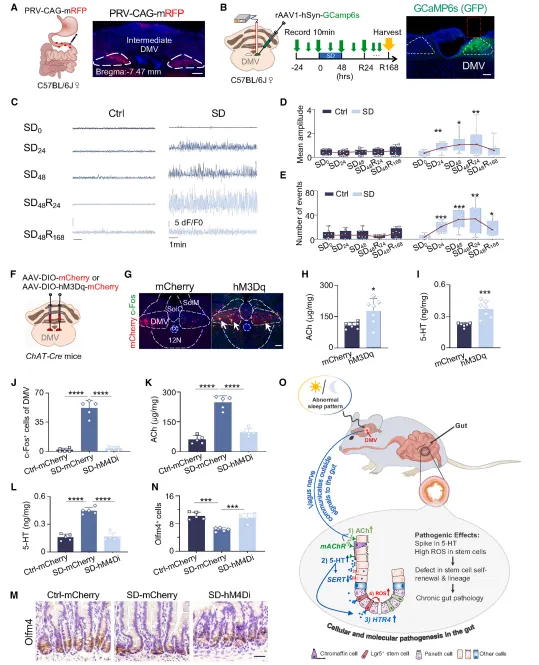
实验观测到,缺觉会让 DMV 神经元长期活跃,时长可达一周。单独激活这个脑区,就能复刻出缺觉带来的肠道问题;缺觉时抑制它,肠道就不会受损伤。这证明 DMV 是整个反应通路的源头(图7)。
图7.DMV 是介导肠道中SD调控的中枢神经系统区域
复现思路:
锁定候选脑区后,先用光纤记录看一下它是否真的被你的干预激活。然后做化学遗传学激活,看能不能复现外周表型;再做抑制实验,看能不能阻断。