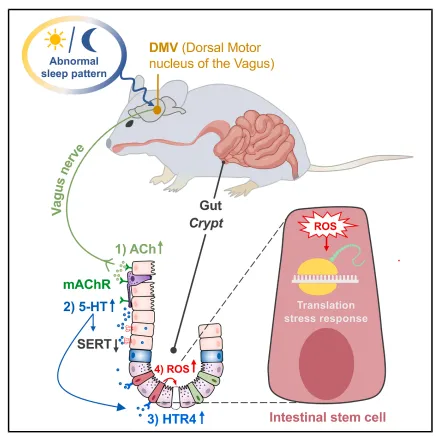
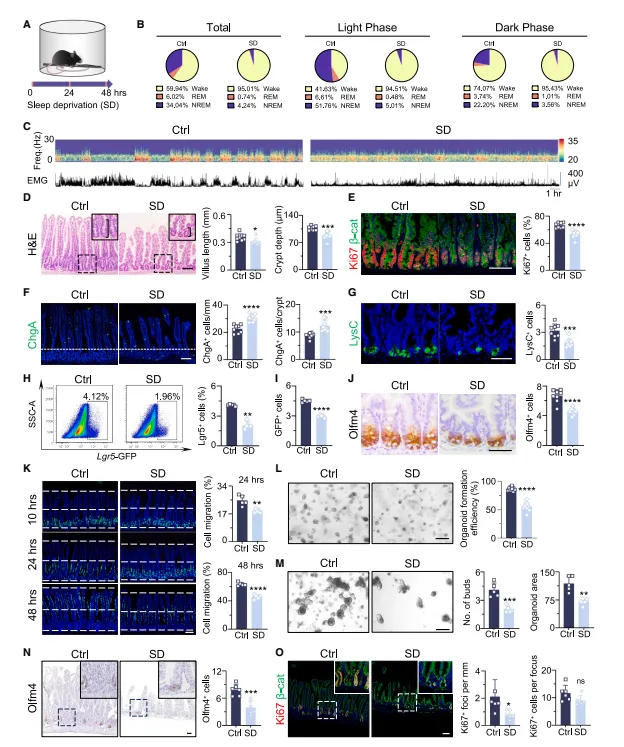
两天睡眠剥夺下来,小鼠小肠绒毛和隐窝明显缩短,增殖细胞减少。我注意到这个表型沿着近端到远端逐渐减弱,空间异质性很清晰,后续取材得盯着近段做(图1)。
图1:短期SD显著损害ISC功能并破坏肠上皮细胞更新
Lgr5和Olfm4阳性细胞数量直接砍半,离体类器官形成能力也跟着下降。这说明睡眠不足不是简单伤黏膜,而是把再生库给掏空了。
复现这个表型不难,机械剥夺两天就能看到近端隐窝缩短。风险在于小鼠应激差异大,建议每组至少8只,否则空间异质性带来的波动会把差异淹掉。
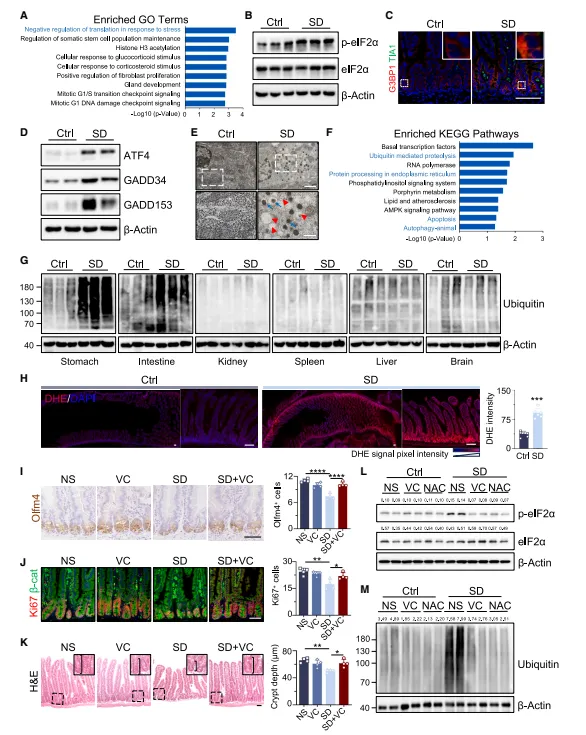
隐窝里磷酸化eIF2α上调,应激颗粒标记物TIA1和G3BP1也多了。翻译应激这条路在肠道干细胞里确实被激活了,算是一个意料之中的损伤信号(图2)。
图2:短期应激损伤通过激活翻译应激反应损害诱导干细胞(ISCs)功能
隐窝蛋白提取效率是瓶颈,建议用激光显微切割富集干细胞区域。全层匀浆做p-eIF2α容易出假阴性,这个靶点对样本新鲜度要求极高。
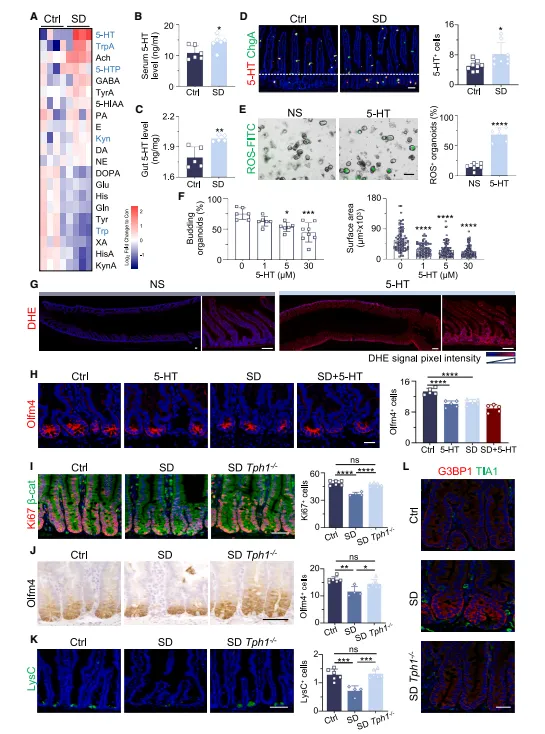
肠道和血清里的5-HT都大幅升高,Tph1敲除后表型就消失了。代谢组筛了一圈,就这个神经递质变化最突出,说明它确实是核心效应分子(图3)。
图3:急性SD可破坏5-HT稳态并诱导肠道氧化应激
代谢组筛差异时要同时检测组织和血清。光看血清可能会漏掉局部变化,我们试过只做血清,5-HT差异并不明显,肠壁才是主战场。
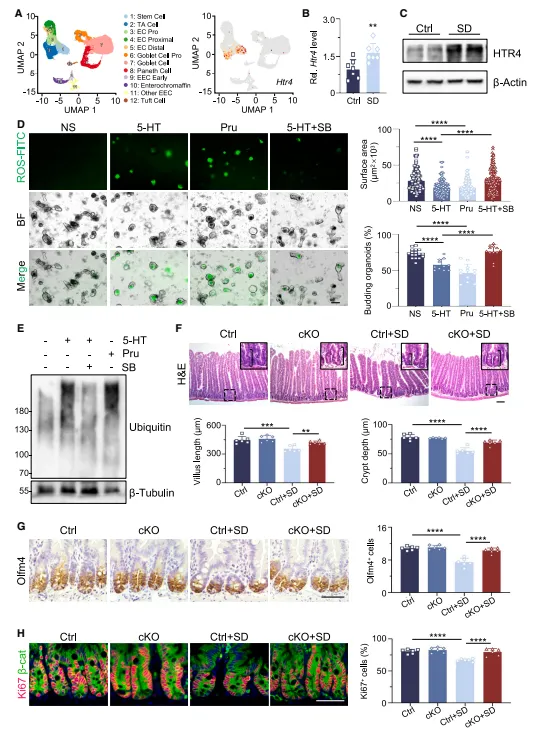
单细胞数据显示Htr4在干细胞里高表达,药物激活它直接复现睡眠剥夺的表型,敲除则保护。受体的特异性这么强,干预窗口比我想的要干净(图4)。
图4:间充质干细胞通过其受体HTR4响应5-羟色胺
单细胞数据里Htr4表达丰度其实不高,做免疫荧光时抗体稀释比要好好摸。另外肠上皮特异性敲除小鼠繁殖周期长,建议提前六个月规划。
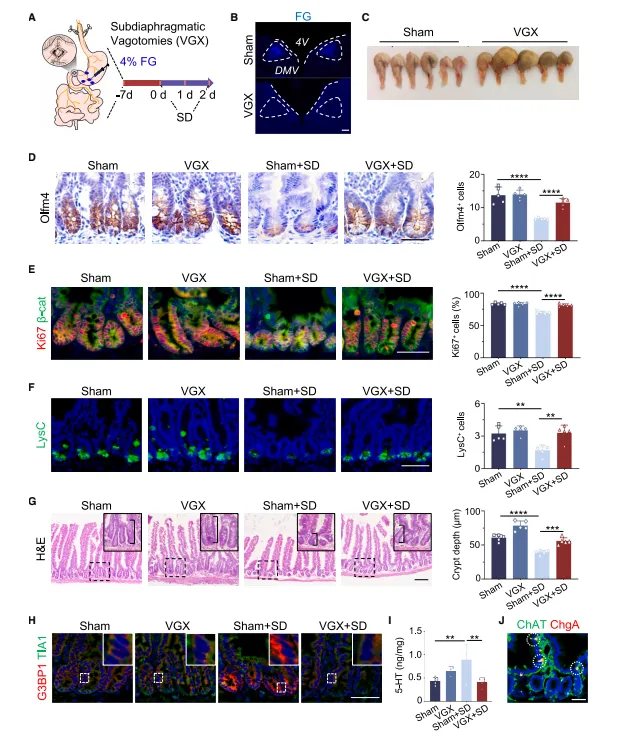
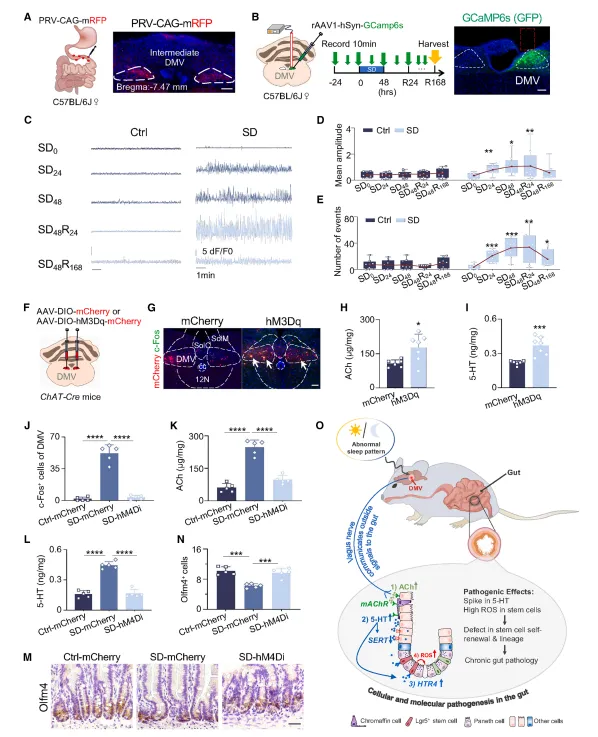
双侧迷走神经切断后,睡眠剥夺引发的肠道损伤被阻断,5-HT也不再升高。反过来化学遗传激活DMV就够出表型了,这套神经回路的因果关系很扎实(图5,图7)。
双侧迷走神经切断术后小鼠存活率只有六成左右,术后前三天需要人工辅助进食。化学遗传的病毒表达周期要卡准,4周刚好,太长会有脱靶。
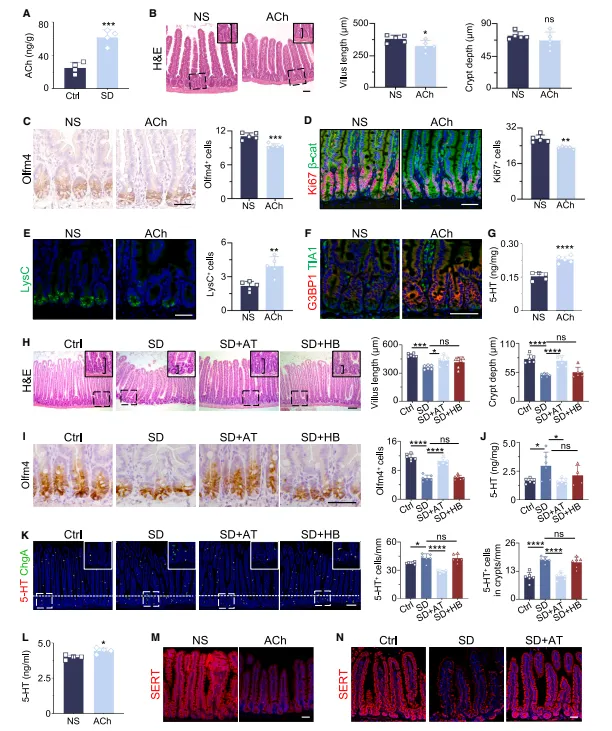
外源给乙酰胆碱就能复刻全部表型,毒蕈碱受体拮抗剂阿托品则完全保护。乙酰胆碱既促进5-HT分泌又抑制其回收,双重拉高局部浓度,这个机制设计得挺巧妙(图6)。
外源给乙酰胆碱代谢太快,我们改成卡巴胆碱效果更稳。阿托品腹腔注射剂量要梯度预试,不同批次小鼠敏感度差异挺大,按体重算不准。