IF 48.5!恭喜郑州大学校长李蓬院士联合北大最新“国自然”研究登顶《Nature》!
磷脂翻转受阻→脂滴在ER腔内堆积→加速脂肪性肝炎。
CLCC1识别膜失衡→招募TMEM41B激活磷脂翻转
如果你也想利用国自然热点+多组学进行生信分析,快关注我们后台咨询吧!生信百事通,助力每一个科研梦想!
发表期刊:Nature
发表时间:2026年2月25日
内质网是脂质合成与分泌的核心枢纽,但其跨膜磷脂转运机制,特别是如何维持脂质稳态以避免代谢性疾病,尚不明确。
综合利用基因编辑、冷冻电子断层扫描、体外功能重建及蛋白质结构预测,结合多组学整合与遗传关联分析,发现ER膜蛋白CLCC1通过促进磷脂翻转酶TMEM41B介导的跨膜磷脂翻转,调控脂蛋白的生物合成与分泌。
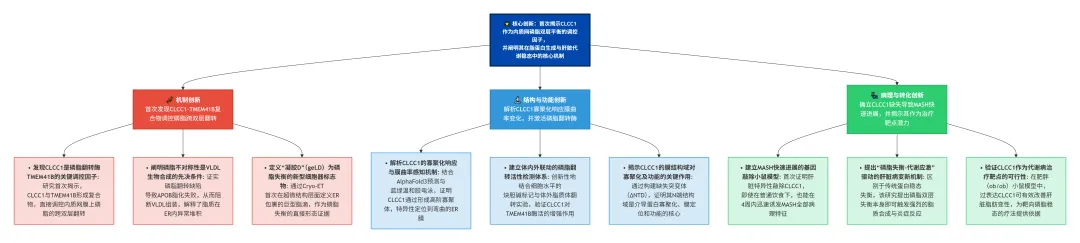
📌三大维度的研究思路框架已整理,点击图片查看。(5分钟获取核心要点)!
《磷脂双层均衡:
CLCC1介导的内质网稳态调控与脂代谢》
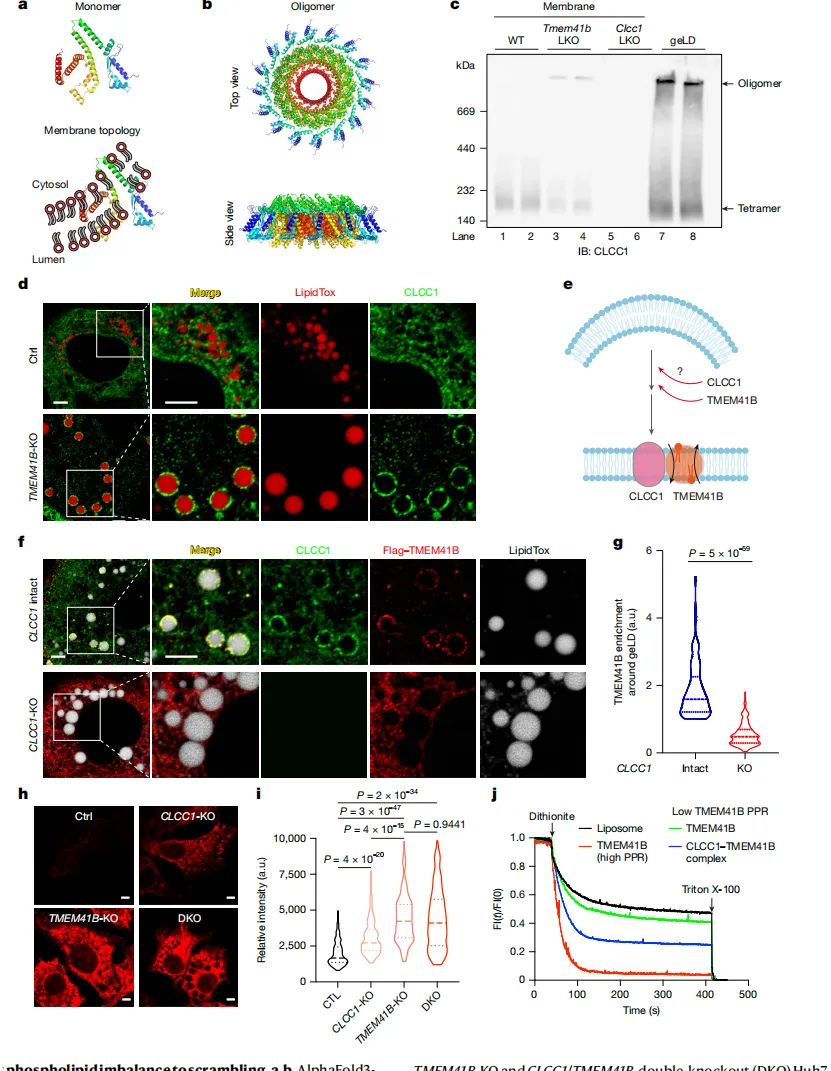
研究团队发现,当肝细胞中关键蛋白TMEM41B缺失后,原本游离的脂滴竟被内质网(ER)的膜整个“吞”了进去,形成一种巨大的“膜包油”结构(gelDs)。
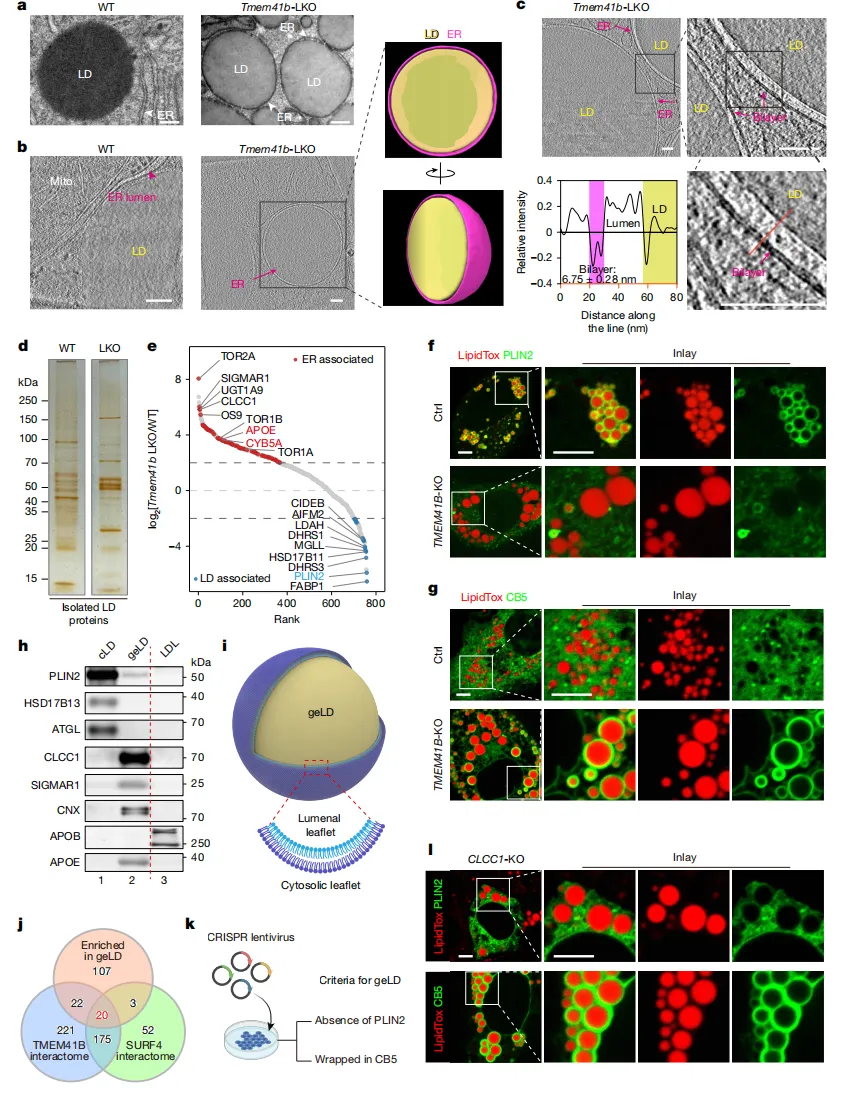
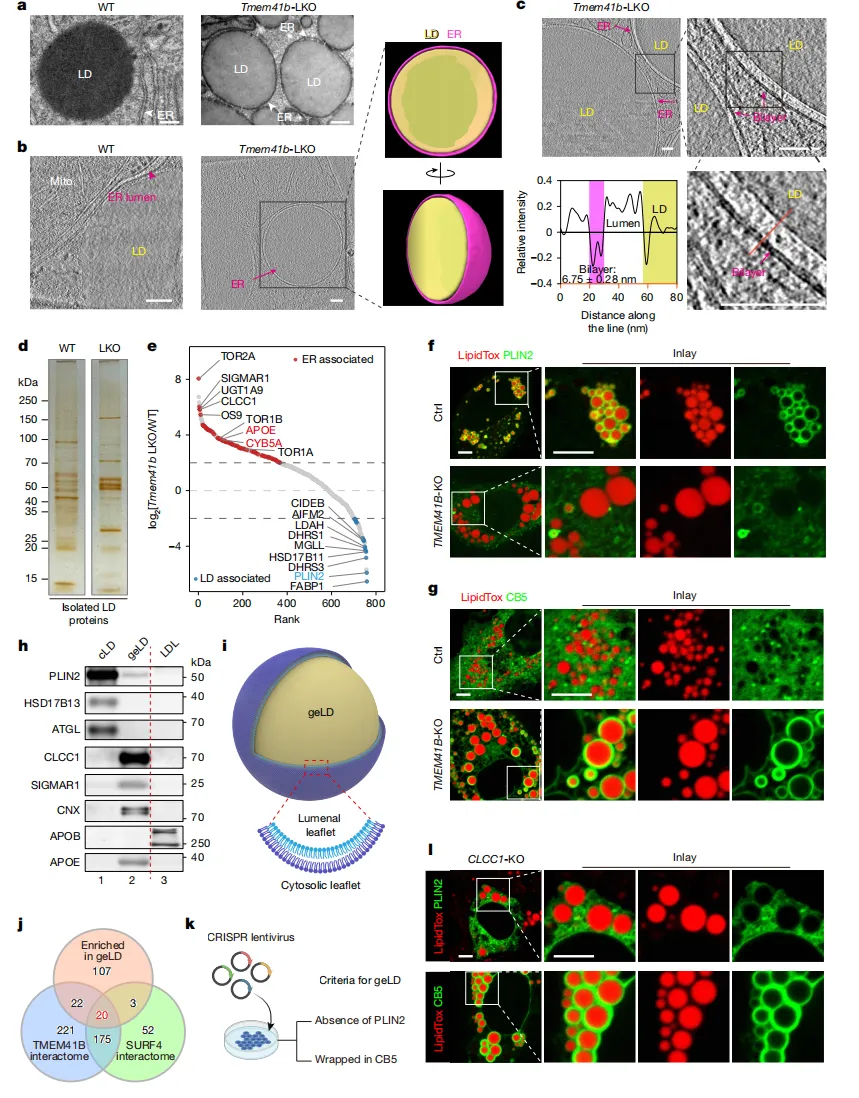
图1.CLCC1或TMEM41B缺失
可触发基因相关疾病(geLDs)
通过三维成像【图1b】可见,正常细胞中脂滴紧挨内质网;而TMEM41B缺陷细胞中,脂滴则被内质网紧紧包裹。
这种异常源于内质网膜内外两层脂质分布不均。细胞通过生成巨大脂滴来缓解膜的压力【图1i】。
对这些脂滴的蛋白质分析发现,它们几乎不含常规脂滴标记蛋白(如PLIN2),反而富集了大量内质网自身蛋白【图1e】。
这种异常脂滴成了宝贵的“探针”。将脂滴富集蛋白与已知调控网络比对后,研究人员锁定了一个此前功能未知的蛋白——CLCC1。
图1.CLCC1或TMEM41B缺失
可触发基因相关疾病(geLDs)
当“敲除”CLCC1基因后,细胞完美复制出与TMEM41B缺失时一模一样的“膜包油”现象【图1l】。
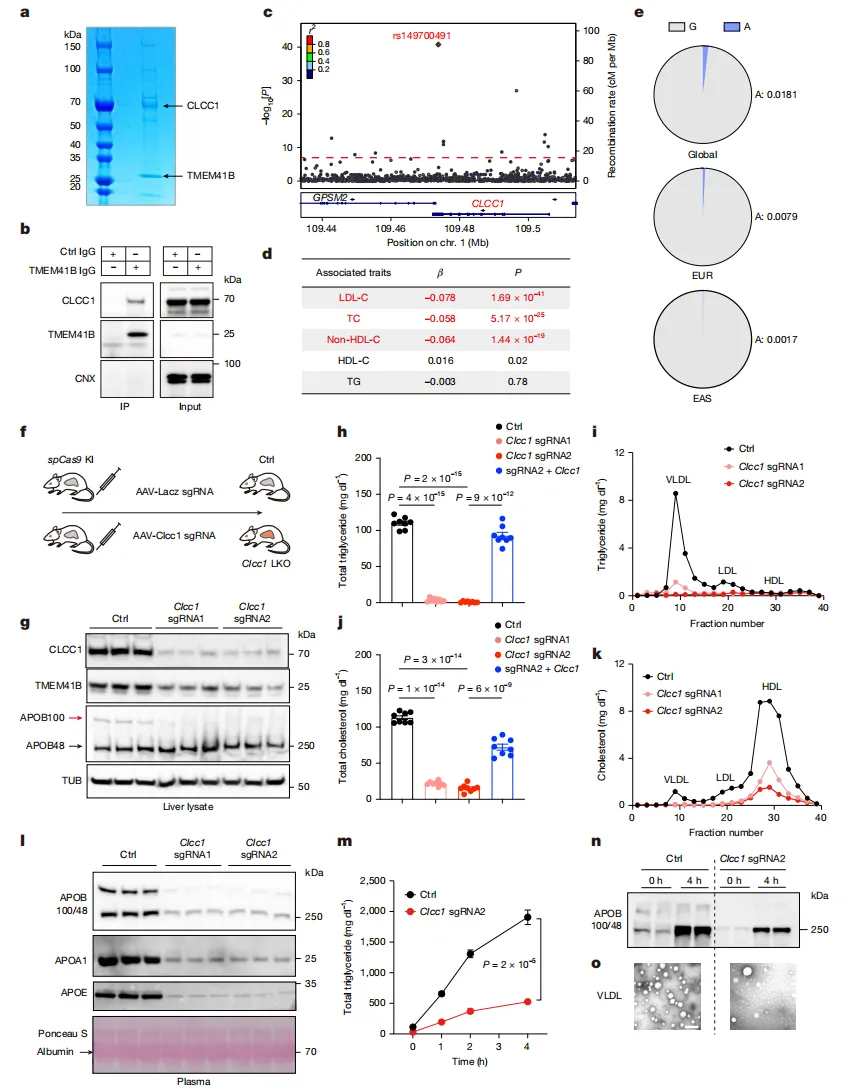
进一步实验证实,CLCC1与TMEM41B是“物理搭档”。人类遗传学数据也显示,CLCC1基因变异与血液中“坏胆固醇”水平显著相关【图2c】。
那么,CLCC1是如何工作的?研究团队发现,它扮演着一个关键的“调度员”角色。
当内质网膜出现脂质失衡时,CLCC1迅速聚集到“不平衡”区域,并“召唤”它的搭档——磷脂翻转酶TMEM41B【图3d】。
体外实验证明,CLCC1本身没有翻转能力,但与TMEM41B“组队”后,却能大幅提升其效率【图3j】。
这解释了为何缺乏CLCC1会导致脂蛋白组装与分泌受阻。
如果这种平衡在肝脏这个“脂质加工厂”中被打破,后果是什么?答案是:一场严重的代谢风暴。
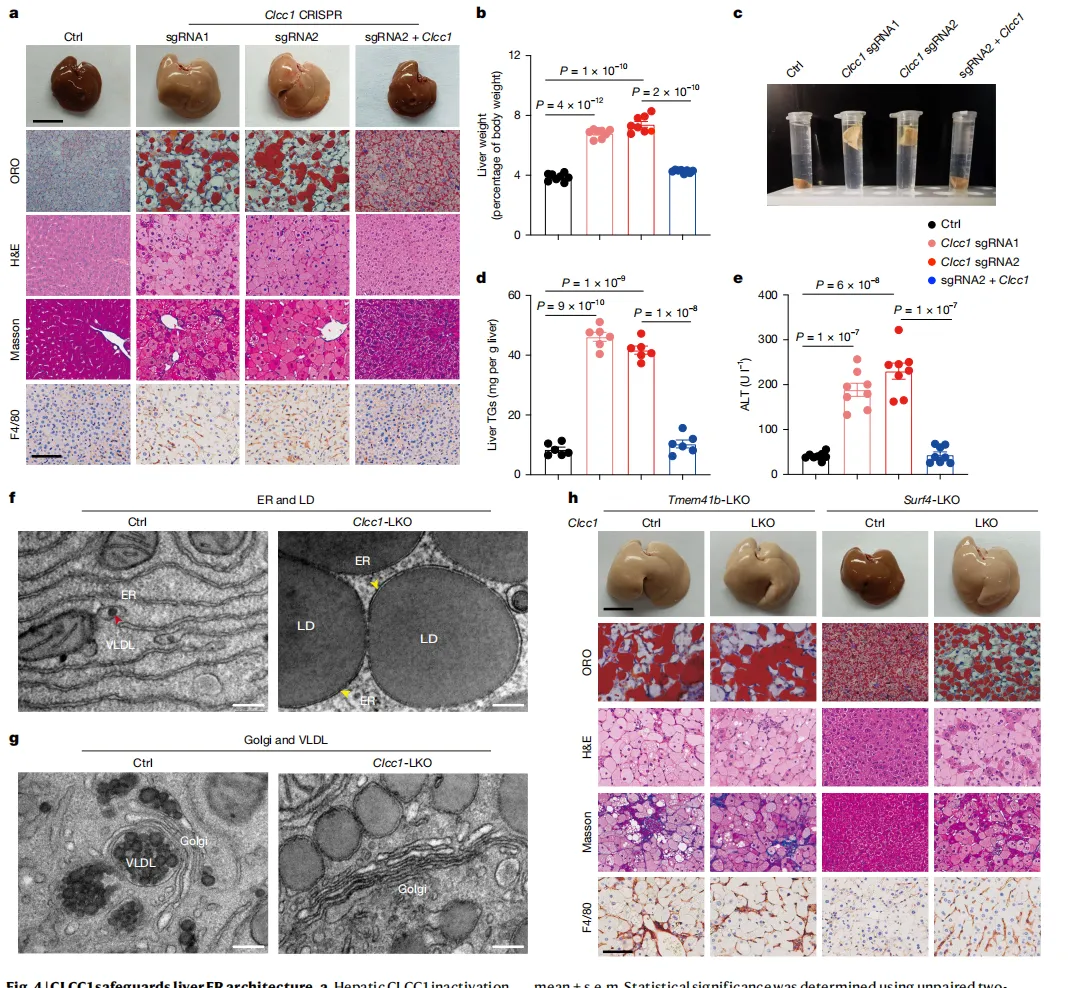
在敲除CLCC1的小鼠模型中,仅4周肝脏就出现严重脂肪性肝炎(MASH),肝脏苍白肿大,甚至能浮在水面上【图4a, c】。
电镜显示,肝细胞被大量凝胶状脂滴充斥,负责运输脂质的“货运火车”VLDL完全消失【图4f, g】。
同时敲掉CLCC1和TMEM41B,病情并未加重【图4h】,证明它们在细胞内处于同一“生产线”。
研究最后,科学家探索了CLCC1在应对代谢应激时的角色。
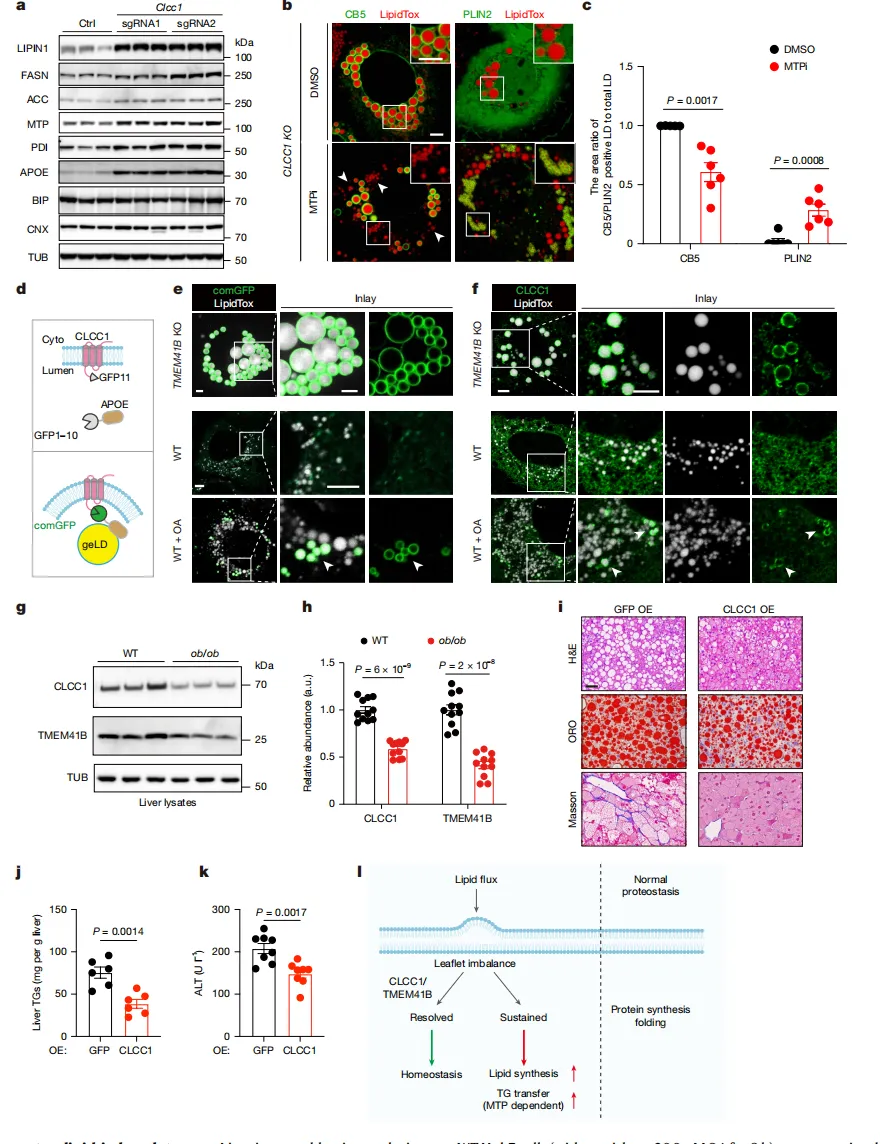
在高脂饮食或肥胖小鼠的肝脏中,CLCC1蛋白水平显著下降【图5g】,暗示它在代谢压力下是“脆弱”的。
当在肥胖小鼠肝脏中“重新补充”CLCC1后,脂肪堆积和肝损伤指标都得到显著改善【图5j, k】。
这一结果不仅印证了CLCC1在脂质稳态中的核心地位,更揭示了它作为潜在“治疗靶点”的价值。
本研究揭示了CLCC1协同TMEM41B维持ER膜磷脂平衡的核心功能,其缺失会引发磷脂失衡与脂质异常堆积,加速代谢相关脂肪性肝炎的病理进程,为干预代谢性疾病提供了新靶点!
有生信分析需求的同学快来联系生信百事通吧!有消息一定及时回复!























 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
